اکسین
اکسینها (Auxin) گروهی از هورمونهای گیاهی هستند که بهخصوص در رشد گیاهان مؤثرند. در ادامه اکسین های طبیعی را با شکل مشاهده می کنید.
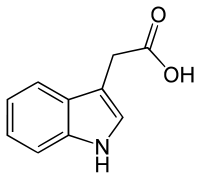
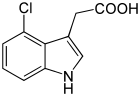
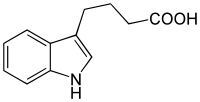
There are three more native — endogenous auxins[1]
All auxins are compounds with aromatic ring and a carboxylic acid group:[2]

تاریخچه اکسین و آزمایشها اولیه

اکسینها اولین هورمون گیاهی کشف شده بودند. چارلز داروین از اولین دانشمندان در تحقیق هورمون گیاهی بود. حاصل مشاهدات او در کتاب «نیروی حرکت در گیاهان» در سال ۱۸۸۰ منتشر شد.
او ابتدا اثراتی از نور در حرکت کولئوپتیلهای زرد روشن شرح داد.
کولئوپتیل یا ساقهپوش یک برگ تخصص یافتهاست که از اولین گرهی که اپیکوتیل را میپوشاند، ناشی میشود که در مرحله حفاظت جوانه گیاهان که از زمین بیرون میآیند .
زمانی که نور یک طرفه به کولئوپتیل میتابد، گیاه در جهت نور خم میشود. اگر رأس کولئوپتیل با فویل آلومینیوم پوشیده شود، به سوی نور یک طرفه خم نمیشود.
ما ا اگر رأس کولئوپتیل در یک طرف پوشیده، یا فقط در قسمتی پایین تر از راس پوشیده شود، نوردهی یک طرفه سبب خمیدگی به سوی نور میشود.
آزمایش داروین پیشنهاد داد که راس کولئوپتیل بافت، مسئول دیدن نور و تولید سیگنالهای انتقالی به قسمت پایین تر کولئوپتیل است جایی که پاسخ فیزیولوژیکی خمیدگی رخ میدهد. او سپس راس کولئوپتیل و باقیمانده کولئوپتیل بدون پوشش را که در معرض نور یک طرفه بود قطع کرد و مطابق نتایج اولیه ازمایشش خمیدگی رخ نداد .
در سال ۱۸۸۵ salkowski ایندول۳-استیک اسیدIAA))را در واسطههای تخمیر کشف کرد. جداسازی محصول یکسان از بافتهای گیاهی پیدا نشد در بافتهای گیاهی برای تقریباً ۵۰ سال.IAA اکسین اصلی دربرگرفته در تعداد زیادی از فرایندهای فیزیولوژیکی در گیاهان است. در سال ۱۹۰۷ fitting اثر ساختمان بریدگی در گیاهان در نور یا تاریکی را مطالعه کرد. نتایجش در جهت فهم این بود که اگر جابجایی سیگنال در یک جهت ویژه گیاه رخ دهد،اما نتایجش ناتمام بود، زیرا سیگنال قابل عبور یا اطراف بریدگی بود.
در سال ۱۹۱۳ Boysen-Jensen آزمایش fitting را اصلاح کردند، به وسیله جاسازی قطعاتی از میکا به عنوان مانع انتقال سیگنال و نشان داد که انتقال اکسین به پایه رخ میدهد در جهت تاریکی گیاه در مقابل جهت در معرض نور یک سوی.
در سال ۱۹۱۸ paalنتایج Boysen-Jensenرا تأیید کرد به وسیلهٔ بریدن راسهای کولئوپتیل در تاریکی، در معرض نور قرار دادن فقط راسها، جابجایی راسهای کولئوپتیل در گیاه بسوی مرکز. نتایج نشان داد که قسمت بدون پوشش کولئوپتیل به جهت دیگر خمیدگی رخ میدهد. Soding دانشمند بعدی بود که تحقیق دربارهٔ اکسین را به وسیله توسعه عقیده paal گسترش داد. او نشان داد که اگر راسها را قطع کنند رشد کاهش مییابد اما اگر آنها بریده میشدند و سپس جابجا میشدند رشد پیوسته رخ میداد.
در سال ۱۹۲۶ یک دانشجوی فارغالتحصیل از هلند که نامش Fritz Went بودیک شرح گزارش را چاپ کرد که او چگونه یک ماده رشد گیاهی را جدا به وسیلهٔ قرار دادن در قطعات اگار زیر راس کولئوپتیل برای یک دوره از زمان کرد سپس آنها را جابجا کردوانها را در ساقههای سربریده جو قرار داد. بعد اگار را جا داد، ساقهها دوباره رشد پیدا کردند. در سال ۱۹۲۸ Went یک روش بری تعیین کمیت ماده رشد این گیاه گسترش داد. نتایجش پیشنهاد کرد که خمیدگی ساقهها متناسب با مقدار ماده رشد در اگار است. این تست، تست خمیدگی جو نامیده شد.
بیشتر دانش اخیر ما از کاربرد اکسین بدست میآید. کار Went یک اثر بزرگی در تحقیق دربارهٔ ماده محرک رشد گیاه داشت. او اغلب با عمل پیراستن به دوره اکسین مهلت داد اما کشف اصلی اکسینها زمانی بود که Kogl وHaagen-Smit ترکیب auxentriolic acid را از ادرار انسان در سال ۱۹۳۱ تصفیه کردند. بعداً Kogl دیگر ترکیبات را را از ادرار جدا کرد که در ساختمان و وظیفه شبیه اکسین A بودند، یکی ایندول ۳- استیک اسید در ابتدا به وسیلهٔ Salkowski در سال ۱۹۸۵ کشف شد. در سال ۱۹۵۴ یک انجمن از فیزیولوژیستهای گیاهی در مشخص کردن گروه اکسینها دائر شد.
Term (دوره) از اکسین یونانی میآید که معنی رشد کردن را میدهد. ترکیبات معمولا اکسینها را مطرح میکنند. اگر آنها به وسیلهٔ گیاه ساخته شوند و مادههایی که فعالیت مشابه به IAA دارند.
بیوسنتز و متابولیسم اکسین
IAAاز نظر شیمیایی به امینو اسید تریپتوفان شبیهاست که معمولاً مولکول را از IAA که اشتقاقی است میپذیرد. ۳ مکانیسم پیشنهاد شده با توضیح این تبدیل: تریپتوفان به ایندول پیروویک اسید با یک واکنش ترانس امیناسیون تبدیل میشود. ایندول پیروویک اسید سپس به ایندول استالدهید به وسیلهٔ یک واکنش دکربوکسیلاسیون تبدیل میشود.
مرحله نهایی شامل اکسیداسیون ایندول استالدهید سبب ایجاد ایندول استیک اسید میشود. ریپتوفان دکربوکسیلاسیون را که سبب ایجاد تریپتامین میشود تحمل میکند. تریپتامین سپس اکسیده میشود و ایندول استالدئید تولید میکند این مولکول اکسیده ایندول استیک اسید تولید میکند. اخیراً در سال ۱۹۹۱ سومین مکانیسم را باز کردند.
IAA میتواند از راه یک مکانیسم مستقل تریپتوفان تولید شود. این مکانیسم بهطور ناچیز فهمیده شدهاست اما استفاده از موتانتهای trp- آزمایش شدهاست. آزمایشهای دیگر نشان دادند که در تعدادی گیاهان این مکانیسم واقعاً مکانیسم مقدم بیوسنتز IAA است. آنزیم مسئول بیوسنتز IAA بیشتر در بافتهای جوان مثل مرستمهای راس جوانه و برگهای در حال رشد و میوهها فعالند.
بافتهای یکسان مکانهایی هستند که بالاترین غلظت IAA را دارند. گیاهان یک جانبه میتوانند مقدار IAA موجود در بافتهایی که در یک زمان معین دقیق هست را به وسیلهٔ کنترل بیوسنتز هورمون کنترل کنند. مکانیسم دیگر کنترل شامل تولید conjugates (درهم آمیختن) که در شرایط ساده هستند مولکولهای که شبیه هورمون هستند اما غیر فعالند. ساختمان conjugates ممکن است یک مکانیسم ذخیره و انتقال هورمون فعال باشد. Conjugates میتوانند از IAA از راه انزیمهای هیدرولاز شکل بگیرند. Conjugatesمیتوانند به سرعت با سیگنالهای محرک محیطی با یک پاسخ هورمونی سریع فعال شوند.
تخریب اکسین روش نهایی کنترل سطح اکسین است.
این فرایند همچنین دو مکانیسم را به صورت زیر پیشنهاد میکند. اکسیداسیون IAA به وسیلهٔ اکسین سبب از دست دادن گروه کربوکسیل و ۳- متیلن اکسیندول میشود محصول تفکیکی اصلی. IAA اکسیداز آنزیمی است که این فعالیت را کاتالیز میکند. Conjugates IAA و اکسینهای ساختگی مثل ۴، ۲ D نمیتواند به وسیلهٔ این فعالیت تخریب شود. C۲ حلقه هتروسیکلیک ممکن است اکسیده شود سبب ایجاد اکسیندول ۳- استیک اسید شود. C3 ممکن است اکسیده شود علاوه بر C۲سبب دی اکسیندول ۳- استیک اسید شود. مکانیسمهایی به وسیلهٔ بیوسنتز و تخریب مولکولهای اکسین رخ میدهند مهم برای کاربرد کشاورزی آینده هستند اطلاعات دربارهٔ متابولیسم اکسین بیشتر مثل هدایت، هدایت به سمت ژنتیک و دستکاری شیمیایی سطح هورمون داخلی سبب رشد مطلوب و تفاوت انواع محصول مهم میشود. سرانجام امکان وجود تنظیم رشد گیاهی بدون استفاده از هربیسیدهای خطرناک و کودها.
وظیفه اکسین
در زیر تعدادی از پاسخهایی که که اکسین سبب میشود:
- تحریک طویل شدن سلول
- تحریک تقسیم سلولی در کامبیوم و در ترکیب با سیتوکینین در کشت بافت
- تحریک تمایز بافت آبکش و بافت چوبی
- تحریک شروع ریشه در برشهای ساقه و پیشرفت ریشه جانبی در کشت بافت
- میانجیگری پاسخ راسی خمیدگی در خمیدگی به جاذبه و نور
- اکسین ذخیره در جوانه راس مانع رشد جوانههای جانبی میشود.
- تأخیر پیری برگ
- جلوگیری یا افزایش ریزش برگ و میوه
- تنظیم و رشد در تعدادی گیاهان
- حرکت یکسان به سمت اکسین ممکن است به وسیلهٔ اثر در حمل و نقل فلوئم در بر بگیرد
- تأخیر رسیدن میوه
- توسعه گلدهی در Bromeliads
- تحریک رشد قسمتهای گل
- گلدهی در گلهای دوپایه
- تحریک تولید اتیلن در غلظت بالا
منابع
- Simon, S; Petrášek, P (2011). "Why plants need more than one type of auxin". Plant Science. 180 (3): 454–460. doi:10.1016/j.plantsci.2010.12.007. PMID 21421392.
- Taiz, L.; Zeiger, E. (1998). Plant Physiology (2nd ed.). Massachusetts: Sinauer Associates.
http://www.plant-hormones.info/auxins.htm
| در ویکیانبار پروندههایی دربارهٔ اکسین موجود است. |