کروماتین
در هنگامی که سلولها در حال تقسیم شدن نیستند، کروماتیدهای آنها به صورت رشتههای نازک و نامنظمی در هسته دیده میشود که به آن کروماتین یا فامینه[1] گفته میشود. کروماتین جنسی یا جسم بار عبارت است از یک جسم کوچک کروماتینی که در مجاورت هسته و چسبیده به غشا به صورت مثلث یا بیضوی دیده میشود و در سلولهای جسمی یا بدنی در جنس ماده و در مرحله ای اینترفاز مشاهده میشوند. درسال ۱۹۴۹ برای اولین بار شخصی به نام «بار» آن را مشاهده کرد واژهٔ کروموزوم به مفهوم جسم رنگیاست، که در سال ۱۸۸۸ بوسیلهٔ والدیر به کار گرفته شد. هماکنون، این واژه برای نامیدن رشتههای رنگپذیر و قابل مشاهده با میکروسکوپهای نوری «LM» به کار میرود که از همانندسازی و نیز به همپیچیدگی و تابیدگی هر رشته کروماتینی اینترفازی در یاختههای یوکاریوتی و رسیدن به ضخامت ۱۰۰۰ تا ۱۴۰۰ نانومتر ایجاد میشود. در پروکاریوتیها نیز مادهٔ ژنی اغلب به حالت یک کروموزوم متراکم میشود. در برخی باکتریها علاوه بر فام تن اصلی که بیشتر ژنها را شامل میشود کروموزوم کوچک دیگری که بهطور معمول آن را پلاسمید مینامند، قابل تشخیص است؛ گرچه تعداد کمی از ژنها بر روی پلاسمید قرار دارند.
اما از آنجا که در بیشتر موارد ژنهای مقاوم به آنتی بیوتیکها بر روی آن جایگزین شدهاند، از نظر پایداری و بقای نسل باکتری اهمیت زیادی دارد. کروماتین در ساختمان کروموزوم به شکل لوپ دیده میشود. لوپها توسط پروتئینهای اتصالی به DNA که مناطق خاصی از آن را تشخیص میدهند، پابرجا میماند. سپس مراحل پیچ خوردگی نهایتاً نوارهایی را که در کروموزومهای متافازی دیده میشود، ایجاد میکند. هر تیپ کروموزومی یک نواربندی اختصاصی را در ارتباط با نوع رنگ آمیزی نشان میدهد. این رنگ آمیزیها منجر به مشخص شدن تعداد و خصوصیات کروموزومهای هرگونه از جانداران میگردد؛ که این خصوصیات تعدادی و مورفولوژیک کروموزومها را کاریوتیپ مینامند.
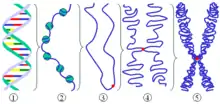
مراحل تبدیل رشته کروماتین به کروموزوم
برای تبدیل یک رشته کروماتینی ۱۰ تا ۳۰ نانومتری به یک کروموزوم، علاوه بر لزوم همانندسازی رشته کروماتین سطوح سازمان یافتگیای را در نظر میگیرند که ضمن آن با دخالت H3، H1 و پروتئینهای غیر هیستونی پیچیدگیها و تابیدگیهای رشتهٔ کروماتین افزایش مییابد، طول آن کم، ضخامت و تراکمش زیاد میشود و به کروموزوم تبدیل میگردد. این سطوح سازمان یافتگی و اغلب به صورت رسیدن از رشتهٔ ۱۰ تا ۳۰ نانومتری به رشته ۹۰ تا ۱۰۰ نانومتری تشکیل رشته ۳۰ تا ۴۰۰ نانومتری و در مراحل بعد با افزایش پیچیدگیها و تابیدگیها، ایجاد رشته ۷۰۰ نانومتری و بالاخره تشکیل کروموزوم دارای دو کروماتید و با ضخامت تا ۱۴۰۰ نانومتر در نظر میگیرند.
اولین مرحلهٔ پیچیدگی و تراکم رشتهٔ کروماتین برای تبدیل به کروموزوم با فسفریلاسیون شدید هیستونهای H3، H1 همراه است. پس از رها شدن DNA از اکتامر هیستونی، با دخالت آنزیمهای مسئول همانندسازی، پیوندهای هیدروژنی بین دو زنجیره گسسته میشود، هر زنجیره مکمل خود را میسازد و به تدریج با ادامهٔ همانندسازی، دو مولکول DNA به وجود میآید که در هر مولکول یک زنجیرهٔ قدیمی و زنجیرهٔ دیگر نوساخت است. بخشهای مختلف این دو مولکول DNA که نظیر یکدیگر هستند به تدریج که همانندسازیشان پایان میپذیرد، با اکتامرهای هیستونی که نیمی از آنها اکتامرهای والدی و نیمی جدید هستند ترکیب میشوند.
بعد از تشکیل ساختمان نوکلئوزومی، دو رشته کروماتین ۱۰ نانومتری و سپس رشتههای ۳۰ نانومتری ایجاد میشوند. هر رشته کروماتین ۳۰ نانومتر سطوح سازمان یافتگی را میگذارند، با مجموعهای از پروتئینهای غیر هیستونی زمینهای یا اسکلتی آمیخته میشود و به یک کروماتید تبدیل میشود. مجموعهٔ دو کروماتید نظیر هم که از محل سانترومر بهم متصلاند کروموزوم متافازی را ایجاد میکنند
کروماتین چیست؟
کروماتین مجموعه ای از DNA و پروتئین است که در سلولهای یوکاریوتی یافت می شود. عملکرد اصلی آن بسته بندی مولکولهای طولانی DNA در ساختارهای فشرده و متراکم تر است. این باعث می شود رشته ها بهم نخورند و همچنین در تقویت DNA در هنگام تقسیم سلول ، جلوگیری از آسیب DNA و تنظیم بیان ژن و همانند سازی DNA نقش مهمی دارد. در حین میتوز و میوز ، کروماتین جداسازی مناسب کروموزومها را در آنافاز تسهیل می کند. شکلهای مشخص کروموزومها که در این مرحله قابل مشاهده هستند ، نتیجه جمع شدن DNA به کروماتین بسیار متراکم است.
اجزای اصلی پروتئین کروماتین هیستون ها هستند که به DNA متصل می شوند و به عنوان "لنگر" هایی عمل می کنند که رشته ها در اطراف آنها زخم می شوند. به طور کلی ، سه سطح سازماندهی کروماتین وجود دارد: DNA به دور پروتئین های هیستون می پیچد و نوکلئوزوم ها و به اصطلاح مهره ها را بر روی یک ساختار رشته ای (euchromatin) تشکیل می دهد. چندین هیستون در یک فیبر 30 نانومتری متشکل از آرایه های نوکلئوزوم در کم حجم ترین شکل (هتروکروماتین) قرار می گیرد.ابرپوشش DNA با سطح بالاتر فیبر 30 نانومتری ، کروموزوم متافاز را تولید می کند (در طی میتوز و میوز). گرچه بسیاری از موجودات از این طرح سازمانی پیروی نمی کنند. به عنوان مثال ، اسپرماتوزوئیدها و گلبول های قرمز پرندگان کروماتین بسته بندی شده محکم تری نسبت به اکثر سلول های یوکاریوتی دارند و پروتوزوآهای trypanosomatid به هیچ وجه کروماتین خود را به کروموزوم های قابل مشاهده متراکم نمی کنند. سلولهای پروکاریوتی ساختارهای کاملاً متفاوتی برای سازماندهی DNA خود دارند (معادل کروموزوم پروکاریوتی ژنوفور نامیده می شود و در منطقه نوکلئوئید قرار دارد).
ساختار کلی شبکه کروماتین بیشتر به مرحله چرخه سلول بستگی دارد. در حین اینترفاز ، کروماتین از نظر ساختاری سست است و اجازه دسترسی به RNA و DNA پلیمرازهایی را می دهد که DNA را رونویسی و تکثیر می کنند. ساختار موضعی کروماتین در طی اینترفاز به ژن های خاص موجود در DNA بستگی دارد. مناطقی از ژنهای حاوی DNA که بصورت فعال رونویسی می شوند ("روشن" می شوند) کمتر فشرده شده و با RNA پلیمرازها در ساختاری معروف به اکروماتین ارتباط نزدیک دارند ، در حالی که مناطق حاوی ژنهای غیرفعال ("خاموش") به طور کلی متراکم تر هستند و با پروتئین های ساختاری در هتروکروماتین. اصلاح اپی ژنتیکی پروتئین های ساختاری در کروماتین از طریق متیلاسیون و استیلاسیون همچنین ساختار محلی کروماتین و در نتیجه بیان ژن را تغییر می دهد. ساختار شبکه های کروماتین در حال حاضر به خوبی درک نشده است و همچنان یک منطقه فعال تحقیق در زمینه زیست شناسی مولکولی است.
منابع
- «معنی فامینه | واژههای مصوّب فرهنگستان». www.vajehyab.com. دریافتشده در ۲۰۲۰-۰۹-۰۷.