جفتباز
جفت باز (base pair)(به اشتباه ، به صورت جفت پایه هم ترجمه شده است) در زیستشناسی مولکولی و ژنتیک، به دو نوکلئوتید با ترکیبات مکمل مخالف روی رشتههای آرانای (RNA) و دیانای (DNA)، که با پیوند هیدروژنی به هم متصل شدهاند، اطلاق میشود. در مدل جفت باز واتسون و کریک برای DNA، آدنین(A) با تیمین(T) و گوانین (G) با سیتوزین(C) جفت باز تشکیل میدهند. در مولکول آرانای، تیمین توسط یوراسیل (U) جایگزین شدهاست. به وجود آمدن مدلهای تناوبی پیوند هیدروژنی، مانند مدلهای جفت باز وابل[1] و جفت باز هوگستین،[2] بخصوص در مولکول آرانای، باعث پیچیدگی و کارآمدن هرچه بیشتر ساختار سوم شدهاست. لازم است ذکر شود که جفت شدن مکانیسمی است که طی آن کدونهای روی مولکولهای mRNA در طی فرایند ترجمه در پروتئین سازی، توسط آنتی کدونهای روی tRNA شناسایی میشوند. برخی از آنزیمهای اتصال دهندهٔ دیانای یا آرانای میتوانند الگوهای جفت باز ساز خاصی را تشکیل دهند که نشانگر مناطق خاص نظارتی از ژن است.
اندازهٔ یک ژن یا تمام ژنوم یک موجود زنده، اغلب با جفتهای باز اندازهگیری میشود. زیرا دیانای معمولاً دورشته ایست؛ بنابراین، تعداد کل جفتهای باز برابر تعداد نوکلئوتیدهای یک رشته میباشد. (به استثناء ساختارهای تکرشتهایهای کدنشدهٔ تلومرها). طول ژنوم هاپلوئید انسانها (با ۲۳ کروموزوم) حدوداً ۳ میلیارد جفت پایه شامل ۲۵٬۰۰۰–۲۰٬۰۰۰ ژن خاص تخمین شدهاست. کیلوبیس (kb)، واحد اندازهگیری در زیستشناسی مولکولی میباشد که برابر ۱۰۰۰ جفت باز دیانای یا آرانای است.
ثبات و پیوند هیدروژنی
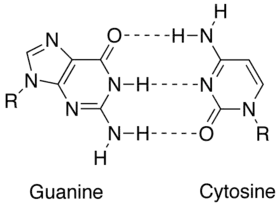

پیوند هیدروژنی یک برهمکنش شیمیایی است که زمینهٔ جفت شدن پایه که در بالا توضیح داده شد، میباشد. درخور تطابق هندسی دهندهها پذیرندهها در پیوند هیدروژنی فقط جفتهای درست و مناسب اجازهٔ تشکیل ساختار پایدار داده میشود. مولکول دیانای دارای محتوای GC زیاد، نسبت به مولکول دیانای دارای محتوای GC کم، استوارتر است. برخلاف باور عموم، پیوندهای هیدروژنی دلیل ثبات دیانای نمیباشد و دلیل این ثبات و پایداری تعاملات پشتهای میباشد.
نوکلئوتیدهای بزرگتر، آدنین و گوانین، اعضای کلاس ساختار شیمیایی دو حلقهای به نام پورین میباشند؛ نوکلئوتیدهای کوچکتر، سیتوزین و تیامین (و یوراسیل)، اعضای کلاس ساختار شیمیایی تک حلقهای که پیریمیدین گفته میشود، هستند. پورینها تنها با پیریمیدینها تکمیل میشوند: جفت پیریمیدین - پیریمیدین از نظر انرژی نامناسب است، زیرا مولکولها برای تشکیل پیوند هیدروژنی زیاد از هم دورند؛ جفت شدن پورین – پورین نیز از نظر انرژی نامناسبند، زیرا این بار مولکولها بیش از حد به هم نزدیکند و همدیگر را دفع میکنند. تنها جفت شدگی ممکن GT و AC میباشد؛ این جفت شدگیها بخاطر عدم تطابق الگوی هیدروژن دهندهها و گیرندههای هیدروژن میباشد. جفت GU، با دو پیوند هیدروژنی، اغلب در آرانای تشکیل میشود.
مولکولهای جفت شدهٔ دیانای و آرانای در دمای اتاق نسبتاً پایدار هستند اما در دمای ذوب دو شاخهٔ نوکلئوتید از هم جدا میشوند و این موضوع به طول مولکول، تعداد جفت شدنهای اشتباه (البته اگر وجود داشته باشد) و تعداد ترکیبات GC دارد. تعداد بیشتر ترکیبات GC باعث بالا رفتن دمای ذوب میشود؛ بنابراین، دور از ذهن نیست که بگوییم ژنوم جانوران سختبومزی[3] مانند گرماپای گرمادوست[4] در اصل غنی از GC میباشد. از طرفی، محدودههایی از ژنوم که لازم است بارها جدا شوند _به عنوان مثال، محدودههای راه انداز رونویسی ژن_ نسبتاً ترکیبات GC کمتری دارند.
ترکیبات GC و دمای ذوب باید به هنگام آمادهسازی مقدمات واکنشهای PCR اندازهگیری شوند.
مثال
توالی دیانای زیر یک الگوی جفت شدن دو رشتهای را نشان میدهد. به صورت قرارداد، رشتهٔ بالایی از سر ۵' به سر ۳' و رشتهٔ پایینی از سر ۳' به سر ۵' میباشد.
- یک دیانای جفت شده:
ATCGATTGAGCTCTAGCG TAGCTAACTCGAGATCGC
- در توالی آرانای زیر جای تیمین با اوراسیل عوض شدهاست:
AUCGAUUGAGCUCUAGCG UAGCUAACUCGAGAUCGC
پانویس
- Wobble base pair
- Hoogsteen base pair
- Extremophile
- Thermus thermophilus
منابع
مشارکتکنندگان ویکیپدیا. «Base pair». در دانشنامهٔ ویکیپدیای انگلیسی، بازبینیشده در ۲۹ ژوئن ۲۰۱۱.