دیانای
دئوکسی ریبو نوکلئیک اسید به اختصار دیاِناِی[1] یا دنا[۱] (به انگلیسی: Deoxyribonucleic acid) (اختصاری DNA)[2] نوعی اسید نوکلئیک است که دارای دستورالعملهای ژنتیکی است که برای کارکرد و توسعهٔ بیولوژیکی موجودات زنده و ویروسها مورد استفاده قرار میگیرد. نقش اصلی مولکول دیانای ذخیرهسازی طولانی مدت اطلاعات ژنتیکی و دستوری است. آزمایشهایی نظیر آزمایش گریفیت و آزمایش ایوری آزمایشهایی انقلابی و سرآغازی در شناسایی و مطالعهٔ دیانای به عنوان ژنوم بودند. تا قبل از سال ۱۹۴۴ و انتشار نتایج آزمایش ایوری اینکه کدام یک از ترکیبهای آلی درون سلول، مادهٔ وراثتی است، مشخص نبود، تا اینکه ایوری ثابت کرد نوکلئیک اسیدها عامل فرایند انتقال صفات هستند. سپس دانشمندان دیگری روی کار آمدند و هر کدام، قسمتی از اطلاعات ما راجع به این مولکول را کشف کردند. روزالیند فرانکلین و موریس ویلکینز با تهیهٔ تصاویری از مولکول دیانای با استفاده از پراش پرتوی ایکس (X) توانستند به ابعاد مولکول و نتایج ارزشمندی راجعبه دیانای دست یابند، از جمله اینکه مولکول دیانای بیش از یک رشته و حالت مارپیچ دارد.

دیانای مولکولی است که دستورهای ژنتیکی مورد استفاده در توسعه و عملکرد تمام موجودات زندهٔ شناخته شده و بسیاری از ویروسها را کدگذاری میکند. دیانای پلیمری است که مونومر آن نوکلئوتیدها هستند؛ یک نوکلئوتید شامل یک گروه فسفات و یک کربوهیدرات پنجکربنه (دئوکسیریبوز) و یک باز آلی است بهطوری که گروه فسفات و باز آلی با پیوند کووالانسی به دو سمت قند متصل هستند؛ و در نهایت نوکلئوتیدها با پیوند فسفو دیاستر به هم متصل شده و درشتمولکول دیانای را پدیدمیآورند (در حالتی که نوکلئوتیدهای دو انتهای رشته هم پیوند برقرار کنند، دیانای حلقوی است)؛ مولکول دیانای از دو رشته پلینوکلئوتیدی تشکیل شدهاست؛ این دو رشته مکمل، ناهمسو و نامحلول (در آب) هستند (دیانای حلقوی قطبیت ندارد اما هر رشته از دیانای خطی دارای قطبیت است). اسیدهای نوکلئیک از سه درشتمولکول اصلی تشکیل شده که برای زندگی همهٔ گونههای شناخته شده ضروری است. دو رشتهٔ پلینوکلئوتیدی از طریق پیوند هیدروژنی بین بازهای آلیشان به هم متصلشده و مولکول دیانای را بهوجود میآورند؛ این دو رشته به دور محوری طولی، مرکزی و فرضی میپیچند و به مولکول دیانای حالت مارپیچ میدهند.
تفاوت دیانای و آرانای
تفاوتها:
- DNA در ذخیره و RNA در انتقال اطلاعات وراثتی و در ساختار ریبوزوم نقش دارد.
- مولکول DNA دو رشتهای در هم تنیده ولی مولکول RNA تکرشتهای است.
- در DNA باز آلی یوراسیل و در RNA باز آلی تیمین شرکت ندارد (U در DNA و T در RNA).
- قند پنجکربنه در DNA دئوکسی ریبوز و در RNA ریبوز است.
- DNA برعکس RNA از هستهٔ سلول خارج نمیشود.
- RNA فاقد ژن میباشد.
شباهتها:
- هر دو پلیمر هستند و از نوکلئوتید تشکیل شدهاند.
- در هر دو نوکلئوتیدهای مقابل با پیوند هیدروژنی و نوکلئوتیدهای مجاور با پیوند فسفو دیاستر به هم متصل میشوند (گاهی نوکلئوتیدهای دو بخش متفاوت از یک رشته آرانای، به هم متصل میشوند).
- نوکلئوتیدهای آزاد (واحدهای سازنده آزاد) هر دو مولکول قبل از اتصال سهفسفات بوده و با اتصال به رشته پلینوکلئوتیدی تکفسفاته میشوند..
بازهای آلی در نوکلئوتیدها
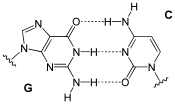
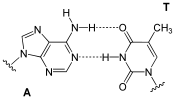
هر نوکلئوتید فقط یک باز آلی نیتروژندار دارد که آن میتواند از نوع آدنین (A) یا گوانین (G) یا سیتوزین (C) یا تیمین (T) باشد. این بازها یا پریمیدین (تکحلقهای) هستند (مانند T و C) یا پورین (دوحلقهای) (A و G).
- مقدار بازهای گوانین و سیتوزین با هم، و تعداد بازهای آدنین و تیمین در یک مولکول دیانای با هم برابر است (اصل چارگاف).
دربارهٔ پیوندی که بین این بازها ایجاد میشود:
- از نوع پیوند بینمولکولی و هیدروژنی است.
- موجب اتصال دو رشتهٔ پلینوکلئوتیدی در مولکول دیانای است.
- به صورت اختصاصی است (یعنی بین آدنین و تیمین و بین گوانین و سیتوزین پیوند به صورت خودکار و طبیعی شکل میگیرد -دو نوکلئوتید مکمل هم هستند-) و بنابراین در هر صورت یک باز تکحلقهای روبروی یک باز دوحلقهای قرار میگیرد که موجب یکسان شدن قطر مولکول میشود.
- بین گوانین و سیتوزین سه پیوند هیدروژنی، و بین آدنین و تیمین دو پیوند برقرار میشود؛ پس پیوند بین گوانین و سیتوزین در برابر فشار و گرما مقاومتر عمل میکند و از شروط پایداری دیانای در این شرایط تعداد بیشتر این دو باز نسبت به آدنین و تیمین است.
- این پیوند بینمولکولی است و نسبت به پیوندهای بیناتمی (مانند کووالانسی) انرژیپیوند کمتری دارد و آسانتر گسسته میشود؛ به همین جهت دیانای را به زیپ تشبیه کردهاند.
تاریخچه کشف ماده وراثتی و ساختار آن

نوکلئیک اسیدها برای اولین بار در زمستان ۱۸۶۹ توسط دانشمند سوئیسی به نام فردریش میشر کشف شد. میشر ترکیبات سفید رنگی را از هستهٔ گلوبولهای سفید انسان و اسپرم ماهی استخراج کرد که مقدار نیتروژن و فسفات در آن باعث شد میشر گروه جدیدی از مواد آلی را با نام نوکلئیک اسیدها بنیانگذاری کند.

سپس فردریک گریفیت در آزمایشی موسوم به آزمایش گریفیت بهطور اتفاقی پی برد صفات و ویژگیها میتوانند از یک باکتری (سلولی) به باکتری (سلولی) دیگری انتقال یابند (اثبات وجود و انتقال مادهٔ وراثتی از سلولی به سلول دیگر).

بعد از آن اسوالد ایوری در سال ۱۹۴۴ نشانداد که مادهٔ وراثتی، نوکلئیکاسید (DNA) است.
در سال ۱۹۵۰ اروین چارگف اثباتکرد نسبت گوانین و سیتوزین با هم و نسبت آدنین و تیمین با هم برابر است (اصل چارگف).
در ۱۹۵۲ روزالیند فرانکلین به همراه موریس ویلکینز با نتایج حاصل از تصاویر گرفتهشده از دیانای با پرتو ایکس نشاندادند دیانای بیش از یک رشته و حالت مارپیچ دارد؛ و در نهایت فرانسیس کریک و جیمز واتسون مدل مولکولی (سهبعدی) دیانای را ارائه دادند؛ که در آن دیانای دو رشتهای بود و مکمل بودن بازها هم مطرحشد.
عملکرد دیانای در سلولها
پیامهای ژنتیکی موجود در مولکول دیانای در نهایت برای مواردی چون ساخت پروتئین و مولکولهای آرانای یا رنا[3] (RNA) در یاخته، مورد استفاده قرار میگیرد. بخشهایی از DNA که اطلاعات ژنتیکی یک صفت را با خود حمل میکنند ژن نامیده میشوند؛ البته دیانای توالیهایی بهنام اینترون نیز دارد که طی فرایند پیرایش از مولکول آرانای حذف میشوند؛ اما نقش زیستی آنها چیست؟
- باعث کاهش آسیبهای مؤثر به دیانای است.
- تنوع در محصولات و گوناگونی آنها.
- تنظیم فرایند رونویسی.
این توالیها در فرایند ترجمه شرکت ندارند.
از لحاظ شیمیایی، دیانای از دو رشتهٔ بلند پلیمری با واحدهای ساختاری بهنام نوکلئوتید تشکیل شدهاست؛ و به یک نردبان مارپیچ تشبیه میشود که ستونهای نردبان گروههای قند و فسفات هستند، و پلههایش را بازهای آلی تشکیل میدهند که با پیوند فسفو دیاستر به هم متصل شدهاند؛ در پیوند فسفو دیاستر قند یک نوکلئوتید به قند نوکلئوتید دیگر متصل میشود.
دو رشتهٔ دیانای با هم موازی و ناهمسو هستند و توالی نوکلئوتیدی خاصی دارند؛ توالی این چهار نوع باز آلی باعث رمزگذاری رشته ژنتیکی میشود که این رمزها برای تعیین توالی اسیدهایآمینه در پروتئین مورد استفاده قرار میگیرد.
طی فرایند رونویسی یک رشته آرانای (با توالی نوکلئوتیدی خاص و مکمل) از روی یک رشته دیانای ساخته میشود؛ به توالیهای سهنوکلئوتیدی آرانایِ پیک، رمز ژنتیکی گویند.
این رمز ژنتیکی برای تعیین توالی اسیدهای آمینه در پروتئین مورد استفاده قرار میگیرد (ترجمه). دیانای در داخل سلول به شکل سازههایی به نام کروموزوم است.
کروموزوم در یوکاریوتها (جانوران، گیاهان، قارچها، آغازیان) در بخشی به نام هستهٔ یاخته قرار دارد، در حالیکه در پروکاریوت (باکتری و آرکیا) در سیتوپلاسم یاخته قرار دارد و جایگاه مشخصی ندارد و به بخشی از غشای پلاسمایی متصل است. در کروموزوم به غیر از مولکول دیانای مجموعهای از پروتئینها که مهمترین آنها هیستونها هستند، وجود دارند که وظیفهٔ فشردهسازی دیانای و تنظیم بیان ژنها را برعهده دارد به گونه ای که دو متر دیانای در سلولی چند میکرومتری جای میگیرد .هیستونها تحت تأثیر عوامل گوناگون از جمله استیلاسیون یا داستیلاسیون هیستونی بسته یا باز میشوند و بدین ترتیب رونویسی از ژنهای ناحیه مربوط به آنها متوقف یا آغاز میشود.
ویژگیها
- دیانای پلیمری است قطبی که از رشتههای تکرار شونده شامل واحدهای سازندهای از جنس نوکلئوتید است. طول رشته زنجیرهای دیانای ۲۲ تا ۲۶ آنگستروم (۲٫۲ تا ۲٫۶ نانومتر) و عرض آن ۳٫۳ آنگستروم یا (۰٫۳۳ نانومتر) است.
- اگرچه هر واحد تکرار شونده دیانای بسیار کوچک است ولی رشتهٔ پلیمری آن ممکن است از میلیونها نوکلئوتید تشکیل شده باشد. برای مثال بزرگترین کروموزوم انسان، کروموزوم شمارهٔ یک دارای طولی به اندازه ۲۲۰ میلیون باز آلی مکمل است.
- دو رشتهٔ سازندهٔ دیانای ساختار در هم پیچیدهای همچون درخت انگور به شکل مارپیچ دارند. یک باز آلی پیوند داده شده به قند نوکلئوزید گفته میشود واگر نوکلئوزید از طریق باز خود به گروه فسفات متصل شود نوکلئوتید تشکیل میشود. اگر چندین نوکلئوتید با یکدیگر پیوند داده شده باشند بهطور مثال در دیانای به آن پلی نکلئوتید گفته میشود.
- رشتههای دیانای از واحدهایی متشکل از قند وگروه فسفات است که به صورت متناوب و تکراری در طول رشته قرار گرفتند.
- قند مورد استفاده در دیانای دئوکسیریبوز که نوعی پنتوز (قند پنج کربنی) است تشکیل شدهاست. قندها توسط گروههای فسفری به یکدیگر پیوند داده شدهاند.
طبقهبندی بازهای نوکلئوتیدی

بازهای نوکلئوتیدی به دو گروه تقسیم میشوند:
- پورینها شامل نوکلئوتید آدنین (A) و گوانین (G) که ترکیبی هتروسیکلیک دارای یک حلقه پنج ضلعی و یک حلقه شش ضلعی هستند.
- پیریمیدینها که یک حلقهٔ شش ضلعی دارند و شامل نوکلئوتید سیتوزین (C) و تیمین (T) هستند. باز آلی یوراسیل (U) هم جزو گروه پیریمیدینها است که معمولاً به جای باز تیمین در ساختار RNA وجود دارد اما در مقایسه با تیمین، در ساختار حلقه خود یک گروه متیل کمتر دارد.
دیانای میتواند در شرایط متفاوت به یکی از حالتهای زیر دیده شود:
| شکلهای مارپیچ DNA | حالت A | حالتB | حالت Z |
|---|---|---|---|
| نسبتهای کلی | کوتاه و پهن | بزرگتر و باریکتر | طویل و باریک |
| ارتفاع به ازای هر جفت باز | ۳/۲آنگستروم | ۲۳/۳آنگستروم | ۸/۳آنگستروم |
| قطر مارپیچ | ۵/۲۵آنگستروم | ۷/۲۳آنگستروم | ۴/۱۸آنگستروم |
| جهت چرخش مارپیچ | راست گرد | راست گرد | چپ گرد |
| خمیدگی باز نسبت به محور مارپیچ | ۱۹+ | ۲/۱- | ۹- |
| متوسط چرخش پروانهای جفت باز | ۱۸+ | ۱۶+ | حدود ۰ |
| موقعیت محور مارپیچ | شیار بزرگ | از میان جفت بازها | شیار کوچک |
| اندازهٔ شیار بزرگ | بسیار باریک با عمق زیاد | پهن و عمق متوسط | پهن شده به روی سطح مارپیچ |
| اندازهٔ شیار کوچک | بسیار پهن ولی کم عمق | باریک و عمق متوسط | بسیار باریک ولی خیلی عمیق |
| صورت بندی پیوند گلیکوزیدی | آنتی | آنتی | آنتی در Cو سین در G |
| جفت باز در هر دور مارپیچ | ۱۱ | ۱۰ | ۱۲ |
حالت B فرم عادی داخل یاخته است.
دیانای یک مارپیچ راست گردان است. اگر دست راست را بالای مولکول دیانای قرار داده بهطوریکه انگشت شصت به سمت بالا و در طول محور بلند مارپیچ باشند و انگشتان شیارها را در مارپیچ دنبال کنند یکی از رشتهها را در جهتی دنبال کنید که انگشت شصت شما اشاره میکند هر جفت باز نسبت به قبلی ۳۶ درجه دور میزند.
جابهجا شدگی باز موقعی رخ میدهد که یک باز از زنجیره خارج شود. این امر باعث متیله شدن باز یا حذف بازهای آسیب دیده میشود. به نظر میرسد زی مایه دخیل در نوترکیبی هم ساخت[4] و همچنین ترمیم دیانای برای یافتن مکانهای هم ساخت یا آسیب دیده شروع به بررسی مولکول دیانای و خارج کردن تک تک بازها میکنند. این عمل انرژی زیادی نیاز ندارد.
چرخش پروانهای حالتی است که باز نسبت به محور بزرگ میچرخد. بهطوریکه ۲ عضو شرکتکننده در یک جفت باز، همیشه بهطور دقیق در یک صفحه نیستند؛ آنها میتوانند یک نظم چرخش پروانهای به خود بگیرند در این نظم ۲ باز در جهت عکس هم حول محور بزرگ جفت باز چرخیده و به جفت باز ویژگی شبیه پروانه میدهد.
واسرشته شدن دیانای حالتی است که وقتی دیانای در دمایی بیش از دمای بدن قرار میگیرد یا در پیاچ بالا قرار دارد حاصل میشود و نیروهای ضعیف بین ۲ رشته از بین رفته و ۲ رشته باز میشود. ۲ رشتهٔ دیانای از آن جایی که به وسیلهٔ نیروهای ضعیف به هم وصل هستند با حرارت دادن محلول تا دمایی بیش از دمای بدن یا تحت شرایط پی هاش بالا میتواند واسرشته شود.
بازسرشته شدن دیانای موقعی ایجاد میشود که ۲ رشتهٔ واسرشته شده در شرایط مناسب دوباره به یکدیگر متصل شوند. یکی از دلایل ناهمگنی ژنی در هوهستهها این است که بعد از وا سرشته شدن با سرعتهای متفاوتی به سمت باز سرشته شدن میروند بعضی بسیار سریع (این قطعهها تعدادشان زیاد است) و بعضی بسیار کند هستند. (این قطعهها تعدادشان کم است)
هیبریدشدگی به معنای این است که ۲ رشته از ۲ منبع مختلف به یکدیگر متصل شوند حتی یکی از رشتهها میتواند آرانای باشد.
افزایش جذب حالتی است که در آن میزان جذب نوری دیانای افزایش مییابد. بیشترین میزان جذب در ۲۶۰ نانومتر دیده میشود که در آن بازها مسئول هستند. با باز سرشته شدن دنا پدیدهٔ کاهش جذب رخ خواهد داد. کاهش جذب به عّلت روی هم قرارگیری باز هاست. اگر دمای محلول دیانای تا دمای آب جوش بالا رود چگالی نوری که جذب میشودبهطور قابل توجهی بالا میرود. نقطهٔ ذوب دیانای که آن را با Tmنشان میدهند دمایی است که در آن دیانای مشابه یخ ذوب میشود و از ساختار نظم دار مارپیچ به ساختار تک رشتهای با نظم کمتر تبدیل میشود. نقطهٔ ذوب بستگی به درصد C:G و قدرت یونی محلول دارد؛ که هرچه بیشتر باشد دما هم افزایش مییابد ذوب شدن پدیدهای تعاونی است.
ابرمارپیچ مثبت و منفی: میزان ابرمارپیچ با اندازهگیری اختلاف بین LK° و LK محاسبه میشود که تفاوت اتصال نامیده میشود. اگر مقدار آن برای یک cccDNA بهطور معنی داری غیر از صفر باشداین دیانای تحت فشار پیچشی قرار دارد؛ و گفته میشود این مولکول دارای ابر مارپیچ منفی است و بر عکس اگر LK>LK° باشد دارای ابر مارپیچ منفی است.
همانندسازی دیانای

قبلاز انجام تقسیمات سلولی بایست مولکولهای دیانای همانندسازی شوند تا اطلاعات وراثتی بدون کموکاست به هر دو سلول دختری انتقال یابند. به فرایندی که طی آن از روی یک مولکول دیانای، مولکول دیانای یکسان و جدید دیگری ایجاد میشود، همانندسازی گویند. در این فرایند ابتدا آنزیمی به نام هلیکاز (helicase) (علت نامگذاری این آنزیم به این نام، به دلیل نوع پیوند بین دو رشته یعنی پیوند هیدروژنی است) دو رشته به هم پیچیده دنا را از هم جدا میکند؛ سپس چند پروتئین بهنام SSBP به دو رشته میچسبند و به آنها اجازه به هم پیوستن دوباره را نمیدهند. در دو طرف هر رشتهٔ اعدادی گذاشته شدهاست که یک طرف '۵ و طرف دیگر '۳ است و در رشتهٔ مقابل برعکس رشته دیگری است؛ مسیر همانند سازی هم همواره از '۵ به '۳ است. آنزیم دیگری به نام DNA پلیمر از 1 (DNA Polymerase I) میآید و همانندسازی (دو برابر شدن DNA) را در یکی از رشتههایی که انتهای '۳ آزاد دارد، انجام میدهد ولی در یکی از رشتهها که انتهای '۳ آزاد ندارد آنزیمی به نام DNA پلیمراز 3 (DNA Polymerase III) میآید و همانندسازی را با روش دیگری انجام میدهد. ابتدا آنزیم دیگری به نام RNA پلیمراز (RNA Polymerase) میآید و قطعههایی از رنا را قرار میدهد و سپس DNA پلیمراز ۳ در کنار این قطعهها مینشیند و همانندسازی را از جای مشخص شدهای ادامه میدهد. سپس دوباره این عمل کمی آن طرفتر یعنی به طرف '۵ انجام میگیرد؛ البته اینجا ۲ مشکل به وجود میآید: ۱. در همانندسازی دنا، قطعههایی از RNA وجود دارد. ۲. بین رشتههای RNA و DNA فاصلههایی وجود دارد که نباید باشند (به قطعات DNA کپی شده در حالتی که '۳باز نیست قطعات اوکازاکی میگویند و واقع آنها DNAهای منقطع هستند).
- برای رفع اشکال اول، آنزیمی به نام RNase H قطعات RNA (هیبرید شده) را برمیدارد و به جای آنها DNA میگذارد که برای این کار نیاز به یون منیزیم دارد.
- برای رفع اشکال دوم، آنزیمی بهنام آنزیم دیانای لیگاز میآید و در فواصل بین DNA جدید که R Nase H به جای RNA گذاشته و دیانایهای منقطع (قطعات اوکازاکی) مینشینند و این فاصلهها را پر میکنند.
همانندسازی در سلولهای یوکاریوتی (جانوران، گیاهان، آغازیان و قارچها) وقتگیرتر و پیچیدهتر از همانندسازی در سلولهای پروکاریوتی است (زیرا هر دیانای در سلول یوکاریوتی چندین برابر دیانای در سلول پروکاریوتی است و تعداد دیانایها نیز بیشتر است)، برای جبران این پیچیدگی سلول یوکاریوتی دو راهکار دارد:
- دیانای یوکاریوتها چندین جایگاه آغاز همانندسازی دارد.
- همانندسازی به صورت دوجهته انجام میشود.
در پروکاریوتها (شامل تمام باکتریها) همانندسازی یکجهته است.[5]
- البته در برخی اوقات همانندسازی در پروکاریوتها نیز دوجهته است و آنها هم میتوانند چندین جایگاه آغاز همانندسازی داشتهباشند (اما غالباً اینطور نیست).
جستارهای وابسته
پانویس
- واژهٔ مصوب فرهنگستان زبان و ادب فارسی، دفتر نخست تا چهارم، ۱۳۷۶ تا ۱۳۸۵
- «DNA». فرهنگستان زبان و ادب فارسی. اکتبر ۲۰۱۸.
- این واژه مطابق با واژگان گردآوری شدهٔ فرهنگستان زبان و ادب فارسی برگردان آرانای است.
- این واژه مطابق با واژگان گردآوری شدهٔ فرهنگستان زبان و ادب فارسی برگردان همولوگ است.
- کتاب اصول زیستشناسی و ژنتیک (دانشگاه آزاد) مؤلفین: سید محمود طباطبایی - محمد رضا عبداللهی
منابع
- Alberts, Bruce; Alexander Johnson, Julian Lewis, Martin Raff, Keith Roberts, and Peter Walters (2002).Molecular Biology of the Cell; Fourth Edition. New York and London: Garland Science. ISBN 0-8153-3218-1
- Mandelkern M, Elias J, Eden D, Crothers D (۱۹۸۱). «The dimensions of DNA in solution". J Mol Biol ۱۵۲ (۱): ۱۵۳–۶۱. doi:۱۰٫۱۰۱۶/۰۰۲۲–۲۸۳۶(۸۱)۹۰۰۹۹–۱. PMID 733890
- Gregory S, et al. (۲۰۰۶). «The DNA sequence and biological annotation of human chromosomeNature ۴۴۱ (۷۰۹۱): ۳۱۵–۲۱. doi:10.1038/nature04727. PMID 16710414
- http://www.chem.qmul.ac.uk/iupac/misc/naabb.html Abbreviations and Symbols for Nucleic Acids, Polynucleotides and their Constituents]IUPAC-IUB Commission on Biochemical Nomenclature (CBN), Accessed 0۳ ژانویه ۲۰۰۶
- Ghosh A, Bansal M (۲۰۰۳). «A glossary of DNA structures from A to Z». Acta Crystallogr D Biol Crystallogr 59 (Pt 4): ۶۲۰–۶. doi:10.1107/S0907444903003251. PMID 12657780
- ژنتیک مولکولی واتسون، نویسنده جیمز واتسون و دیگران، گروه مترجمین خانهٔ زیستشناسی، ناشر خانهٔ زیستشناسی، چاپ اول
- Ponnuswamy P, Gromiha M (۱۹۹۴). «On the conformational stability of oligonucleotide duplexes and tRNA molecules". J Theor Biol ۱۶۹ (۴): ۴۱۹–۳۲. doi:10.1006/jtbi.1994.1163. PMID 7526075
- Clausen-Schaumann H, Rief M, Tolksdorf C, Gaub H (۲۰۰۰)"Mechanical stability of single DNA molecules Biophys J ۷۸ (۴): ۱۹۹۷–۲۰۰۷. PMID 10733978
- Chalikian T, Völker J, Plum G, Breslauer K (۱۹۹۹)A more unified picture for the thermodynamics of nucleic acid duplex melting: a characterization by calorimetric and volumetric techniques Proc Natl Acad Sci USA ۹۶ (۱۴): ۷۸۵۳–۸. doi:10.1073/pnas.96.14.7853. PMID 10393911
- زیستشناسی و آزمایشگاه ۲، سال سوم متوسطه، نظری (رشتهٔ علوم تجربی)، چاپ دوازدهم: ۱۳۹۱، دفتر برنامهریزی و تألیف کتب درسی وزارت آموزش و پرورش. شابک:۹-۰۹۸۰-۰۵-۹۶۴
انواع نوکلئیک اسیدها | |||||||
|---|---|---|---|---|---|---|---|
| مؤلفات سازنده | |||||||
| RNA (RNA کدکننده و غیرکدکننده) |
| ||||||
| DNA |
| ||||||
| آنالوگ |
| ||||||
| حاملان کلونینگ | |||||||