دیانای میتوکندریایی
دیانای میتوکندریایی (mtDNA) نوعی دنا است که در میتوکندری سلولهای یوکاریوتی یافت میشود. کار میتوکندری تبدیل انرژی شیمیایی غذا به آدنوزین تری فسفات یعنی صورتی از انرژی است که برای سلول قابل استفاده باشد.
رونویسی
mtDNA به وسیلهٔ پلیمراز گاما رونویسی میشود که به وسیلهٔ ژنوم هستهای کد میشود. رونویسی DNA میتوکندریایی الزاماً با تقسیم میتوکندری همراه نیست به همین دلیل ممکن است در یک میتوکندری چندین نسخه از ژنوم بهطور جداگانه موجود باشد که به ان کنکاتامر (concatamer) میگویند.[2]
خاستگاه
به نظر میرسد DNA میتوکندریایی و هسته دارای ویژگیهای تکاملی متفاوتی باشند. mtDNAها از ژنومهای حلقوی باکتریهایی که توسط اجداد اولیهٔ سلولهای یوکاریوتی امروزی در بر گرفته شدهاند مشتق شدهاند. این نظریه به نظریهٔ اندوسیمبیوتیک (endosymbiotic theory) معروف است. بهطور تقریبی هر میتوکندری شامل ۲–۱۰ کپی از mtDNA میباشد.[3] در سلولهای ارگانیسمهای موجود اکثریت بسیار بزرگی از پروتئینهای موجود در میتوکندری به وسیلهٔ DNA هسته کد میشوند اما تصور میشود ژنهای برخی از آنها دارای ریشهٔ باکتریایی هستند که در طی تکامل به سلولهای یوکاریوتی منتقل شدهاند.
وراثت میتوکندریایی
در اغلب پر سلولیها mtDNA از مادر به ارث میرسد. مکانیزمهای این توارث عبارتست از یک رقیقسازی ساده (یک سلول تخم شامل ۱۰۰هزار تا یک میلیون مولکول mtDNA است در صورتی که یک اسپرم تنها شامل ۱۰۰ تا هزار عدد از انهاست)، کاهش mtDNA اسپرمی در یک تخم بارور شده و حداقل در تعداد کمی از ارگانیسمها ناکامی mtDNAهای اسپرمی در ورود به تخم. فارغ از اینکه چه مکانیسمی مؤثر واقع شود این الگوی تک والدی بودن mtDNA در اکثر جانوران، گیاهان و قارچها دیده شدهاست.
وراثت ماده
در تولید مثل جنسیتی میتوکندری بهطور انحصاری از مادر به ارث میرسد. میتوکندری موجود در اسپرم پستانداران معمولاً پس از لقاح توسط سلول تخم نابود میشود. علاوه بر این بیشتر میتوکندری در پایهٔ دم اسپرم حضور دارد که به منظور به پیش راندن اسپرم استفاده میشود و گاهی اوقات دم در ضمن فرایند لقاح نابود میشود. در ۱۹۹۹ این نتیجه بدست آمد که میتوکندری اسپرمی والدی (parental) توسط اوبیکوتین (ubiquitin) علامتگذاری میشود تا در آینده برای انهدام درون جنینی انتخاب شود.[4] برخی تکنیکهای لقاح مصنوعی به ویژه تزریق یک اسپرم به درون یک oocyte ممکن است با این فرایند تداخل کند. این حقیقت که mtDNA از طریق مادری به ارث میرسد محققان را قادر میسازد تا سلسلهٔ نسل مادری را در طی زمان ردیابی کنند (به طریق مشابه DNAهای کروموزوم Y که از طریق پدری به ارث میرسد برای دنبال کردن سلسلهٔ نسل پدری بکار میرود). این کار در انسانها به وسیلهٔ آنالیز توالی یک یا چند بخش از نواحی کنترل (HVR1 یا HVR2) بس متغیر (hypervariable)، مولکول mtDNA و در قالب یک تست DNA نسب شناسانه انجام میشود.HVR1 از حدود ۴۴۰ جفت باز (Base pair) تشکیل شدهاست. این ۴۴۰ جفت باز با نواحی کنترل افراد دیگر (اشخاص دیگر یا منابع موجود در دیتابیس) به منظور مشخص کردن شجرهٔ مادری مقایسه میشوند. vila et al نتایج تحقیقات نسبشناسی نوعی سگ بومی را تا درندگان منتشر کردهاست.[5] مفهوم حوای میتوکندریایی نیز براساس تحلیلی مشابه بنا شد تا از طریق ردیابی نسل در طول زمان ریشهٔ اولیهٔ بشر را کشف کند. از آنجا که mtDNA بهطور کامل بکر نمانده و نرخ جهش سریعی دارد، میتواند برای بررسی روابط تکاملی ارگانیسمها مفید واقع شود. در واقع میتوان توالی mtDNAها را در گونههای مختلف مشخص کرد و با مقایسهٔ آنها یک درخت تکاملی ترسیم کرد. از آنجا که mtDNA از مادر به فرزند منتقل میشود میتوان از ان به عنوان ابزاری مفید در تحقیقات نسبشناسی برای پیدا کردن اجداد مادری فرد استفاده کرد.
وراثت نر
مشاهده شده که میتوکندری در بعضی از گونهها مانند صدفها میتواند از پدر به ارث برسد.[6][7] همچنین وراثت پدری میتوکندری در برخی حشرات چون مگس میوه،[8] زنبور عسل[9] و برخی جیرجیرک ها[10] نیز گزارش شدهاست. شواهد حاکی از این هستند که توارث پدری میتوکندری در میان پستانداران بسیار نادر است. بهطور مشخص نتایج ثبت شدهای در مورد موشها موجودند که میتوکندریهای پدری پس زده شدهاند.[11][12] این مسئله در میان گوسفندان[13] و همچنین گاوهای شبیهسازی شده یافت شده است[14] و همچنین در یک مورد خاص انسانی.[15]
ساختار
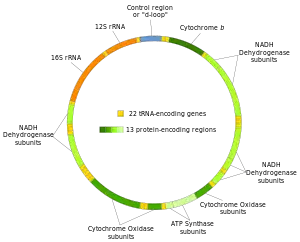
در انسانها و احتمالاً بهطور کلی در همهٔ پرسلولیها در هر سلول بین ۱۰۰ تا ۱۰۰۰۰ کپی متفاوت از mtDNA موجود است (سلول تخم و اسپرم استثنا هستند). در پستانداران هر مولکول mtDNA حلقوی ۲رشتهای شامل ۱۵۰۰۰–۱۷۰۰۰ جفت باز میباشد. ۲رشتهٔ mtDNA بر اساس محتوی هسته ایشان با هم متفاوت میشوند بهطوریکه رشتهٔ با گوانین بیشتر رشتهٔ سنگین تر و رشته با سیتوزین بیشتر با عنوان رشتهٔ سبک یاد میشوند. در یک مجموع ۳۷ ژنی، رشتهٔ سنگین ۲۸ ژن و رشتهٔ سبک ۹ ژن را کد میکند. در این ۳۷ ژن ۱۳ تا برای پروتئینها (پلی پپتیدها)، ۲۲تا برای tRNAها و دو تا برای زیربخشهای کوچک و بزرگ rRNAها هستند. این الگو همچنین در میان اکثر پرسلولیها مشاهده شدهاست. با این وجود در برخی از موارد یکی یا چند مورد از ۳۷ ژن غایب اند و محدودهٔ اندازهٔ mtDNAها بزرگتر است. حتی در میان گیاهان و قارچها تغییرات بزرگتری در محتوای ژنتیکی و اندازهٔ mtDNAها مشاهده شدهاست. همچنین به نظر میرسد یک زیرمجموعه از ژنها وجود دارد که در همهٔ سلولهای یوکاریوتی حضور دارد (به استثنای بعضی موارد که بهطور کلی میتوکندری ندارند). برخی گونههای گیاهی مقادیر بسیار زیادی mtDNA دارند (۲۵۰۰۰۰۰ جفت باز در مولکول mtDNA) با این وجود به طرز غیرمنتظرهای حتی این mtDNAهای بزرگ تعداد و انواع برابری ژن در مقایسه با گیاهان با mtDNA کوچکتر دارند.[16]
ژنها
زنجیرهٔ انتقال
ژنوم میتوکندریایی شامل ۱۳ ژن کدکنندهٔ پروتئین است. بسیاری از این ژنها زنجیرهٔ انتقال را کد میکنند.
| دستهها | ژنها |
| NADH dehydrogenase (complex I) | MT-ND1, MT-ND2, MT-ND3, MT-ND4, MT-ND4L, MT-ND5, MT-ND6 |
| Coenzyme Q - cytochrome c reductase/Cytochrome b (complex III) | MT-CYB |
| سیتوکروم اکسیداز (complex IV) | MT-CO1 , MT-CO2 , MT-CO3 |
| ایتیپی سینتیز | MT-ATP6, MT-ATP8 |
rRNA
rRNA میتوکندریایی توسط MT-RNR1 (12s) و MT-RNR2 (16s) کد میشود.
tRNA
ژنهای زیر tRNA را کد میکنند:
| اسید آمینه | 3-Letter | 1-Letter | MT DNA |
|---|---|---|---|
| آلانین | Ala | A | MT-TA |
| آرژینین | Arg | R | MT-TR |
| آسپاراژین | Asn | N | MT-TN |
| اسید آسپارتیک | Asp | D | MT-TD |
| Cysteine | Cys | C | MT-TC |
| اسید گلوتامیک | Glu | E | MT-TE |
| گلوتامین | Gln | Q | MT-TQ |
| گلیسین | Gly | G | MT-TG |
| هیستیدین | His | H | MT-TH |
| ایزولوسین | Ile | I | MT-TI |
| لوسین | Leu | L | MT-TL1, MT-TL2 |
| لیزین | Lys | K | MT-TK |
| متیونین | Met | M | MT-TM |
| فنیلآلانین | Phe | F | MT-TF |
| پرولین | Pro | P | MT-TP |
| سرین | Ser | S | MT-TS1, MT-TS2 |
| ترئونین | Thr | T | MT-TT |
| تریپتوفان | Trp | W | MT-TW |
| تیروزین | Tyr | Y | MT-TY |
| والین | Val | V | MT-TV |
جهش
بیماریهای ژنتیکی
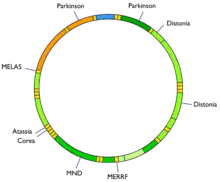
جهش در mtDNA میتواند منجر به شماری از بیماریهای ژنتیکی مانند exercise intolerance و سندروم کرنز-سایر (kss) شود که میتواند سبب کاهش کارکرد قلب، چشمها و حرکات ماهیچه شود. برخی شواهد حاکی از انند که این نقصها نقش اساسی در فرایند پیری دارند.[17]
کاربرد در شناسایی
در انسانها mtDNA، تعداد ۱۶۵۶۹ بلوک سازندهٔ DNA (جفتهای باز) را شامل میشود[18] که نمایش دهندهٔ بخشی از مجموع DNA موجود در سلول است. برخلاف DNA هسته که از هر دو والد به ارث میرسد و در ان ژنها در طی فرایند نوترکیبی چینشی تازه مییابند در mtDNA معمولاً از والد به فرزند تغییری مشاهده نمیشود. با این وجود mtDNA نیز دچار نوترکیبی میشود و این کار را با کپیهایی از خودش در یک میتوکندری واحد انجام میدهد. به این دلیل و نیز به دلیل اینکه در حیوانات mtDNA نرخ جهش بالاتری نسبت به DNA هسته دارد[19] مولکول mtDNA ابزاری قوی برای ردیابی نسلها از طریق مادری است و از این طریق برای جستجوی اجداد بسیاری از گونهها در طول صدها نسل بکار میرود. mtDNA انسان همچنین میتواند برای شناسایی افراد بکار رود.[20] مراکز پزشکی قانونی گاهی اوقات مقایسههای mtDNA را برای شناسایی بقایای انسانی و به ویژه شناسایی بقایای اسکلتهای قدیمی به کار میبرند. گرچه mtDNA برخلاف DNA هسته تنها مختص به یک شخص نیست اما میتوان با استفاده مشترک از ان و شواهد دیگر (مانند شواهد ریختشناسی، شواهد مکانی و...) عمل شناسایی را انجام داد. همچنین به عنوان گواهی سلبی نیز عمل میکند.[21] بسیاری از محققان معتقدند mtDNA نسبت به DNA هستهای ابزار بهتری برای شناسایی بقایای اسکلتهای قدیمی است زیرا به دلیل وجود کپیهای زیاد از mtDNA در سلول احتمال بدست آوردن یک نمونهٔ مفید افزایش مییابد و به این خاطر که یک همسانی با یک ارتباط زنده بسیار محتمل است حتی اگر فواصل نسلی مادری زیادی آنها را از هم جدا کند. جسد یک تبهکار معروف آمریکایی از همین طریق و به وسیلهٔ مقایسهٔ mtDNA او با یکی از اعقاب مسیر دختری وی مشخص شد.[22] همینطور تشخیص هویت برخی از اعضای خاندان سلطنتی روسیه به وسیله مقایسه با اقوام مادری آنها.[23] میزان کم جمعیت مؤثر و نرخ جهش سریع (در حیوانات) mtDNA را برای یافتن روابط ژنتیکی بین افراد و گروهها در یک گونهٔ خاص و همچنین شناسایی و دستهبندی فیلوژنی بین گونههای مختلف ابزاری مناسب ساختهاست البته مشروط به اینکه این گونهها ارتباطشان از هم خیلی دور نباشد. این کار به این شکل صورت میگیرد که ابتدا توالی mtDNA افراد یا گونههای مختلف مشخص میشود و آنگاه بر حسب رابطهای که این توالیها با هم دارند شبکهای ایجاد میشود که درخت فیلوژنتیک نام دارد.
تاریخچه
mtDNA در ۱۹۶۰ توسط مارگریت ام.کی. ناس و سیلوان ام. کی ناس و به وسیلهٔ میکروسکوپ الکترونی[24] و همچنین توسط الن هاسلبروز، هانس تاپی و گاتفرید شاتز به وسیلهٔ ازمایشهای بیوشیمیایی روی بخشهای به شدت خالص شدهٔ میتوکندری کشف گردید.[25]
جستارهای وابسته
منابع
- Iborra FJ, Kimura H, Cook PR (2004). "The functional organization of mitochondrial genomes in human cells". BMC Biol. 2: 9. doi:10.1186/1741-7007-2-9. PMC 425603. PMID 15157274.
- مهدوی، موسوی، اردستانی، صادقی زاده، زیستشناسی سلولی مولکولی و مهندسی ژنتیک/ خانه زیستشناسی
- Wiesner RJ, Ruegg JC, Morano I (1992). «Counting target molecules by exponential polymerase chain reaction, copy number of mitochondrial DNA in rat tissues». Biochim Biophys Acta. 183 (2): 553–559. PMID 1550563
- Sutovsky, P. , et al. (Nov. 25, 1999). «Ubiquitin tag for sperm mitochondria». Nature 402 (6760): 371–372. doi:10.1038/46466
- Vilà C, Savolainen P, Maldonado JE, and Amorin IR (13 June 1997). «Multiple and Ancient Origins of the Domestic Dog». Science 276 (5319): 1687–1689. doi:10.1126/science.276.5319.1687. PMID 9180076
- Hoeh WR, Blakley KH, Brown WM (1991). «Heteroplasmy suggests limited biparental inheritance of Mytilus mitochondrial DNA». Science 251 (5000): 1488–1490. doi:10.1126/science.1672472. PMID 1672472.
- Penman, Danny (23 August 2002). «Mitochondria can be inherited from both parents». NewScientist.com. Retrieved 2008-02-05.
- Kondo R, Matsuura ET, Chigusa SI (1992). «Further observation of paternal transmission of Drosophila mitochondrial DNA by PCR selective amplification method,». Genet. Res. 59 (2): 81–4. doi:10.1017/S0016672300030287. PMID 1628820.
- Meusel MS, Moritz RF (1993). «Transfer of paternal mitochondrial DNA during fertilization of honeybee (Apis mellifera L.) eggs». Curr. Genet. 24 (6): 539–43. doi:10.1007/BF00351719. PMID 8299176.
- Fontaine, KM, Cooley, JR, Simon, C (2007). «Evidence for paternal leakage in hybrid periodical cicadas (Hemiptera: Magicicada spp.)». PLoS One. 9 (9): e892. doi:10.1371/journal.pone.0000892. PMC 1963320. PMID 17849021.
- Gyllensten U, Wharton D, Josefsson A, Wilson AC (1991). «Paternal inheritance of mitochondrial DNA in mice». Nature 352 (6332): 255–7. doi:10.1038/352255a0. PMID 1857422.
- Shitara H, Hayashi JI, Takahama S, Kaneda H, Yonekawa H (1998). «Maternal inheritance of mouse mtDNA in interspecific hybrids: segregation of the leaked paternal mtDNA followed by the prevention of subsequent paternal leakage». Genetics 148 (2): 851–7. PMC 1459812. PMID 9504930.
- Zhao X, Li N, Guo W, et al. (2004). «Further evidence for paternal inheritance of mitochondrial DNA in the sheep (Ovis aries)». Heredity 93 (4): 399–403. doi:10.1038/sj.hdy.6800516. PMID 15266295.
- Steinborn R, Zakhartchenko V, Jelyazkov J, et al. (1998). «Composition of parental mitochondrial DNA in cloned bovine embryos». FEBS Lett. 426 (3): 352–6. doi:10.1016/S0014-5793(98)00350-0. PMID 9600265.
- Schwartz M, Vissing J (2002). «Paternal inheritance of mitochondrial DNA». N. Engl. J. Med. 347 (8): 576–80. doi:10.1056/NEJMoa020350. PMID 12192017.
- Ward BL, Anderson RS, Bendich AJ (September 1981). «The mitochondrial genome is large and variable in a family of plants (cucurbitaceae)». Cell 25 (3): 793–803. doi:10.1016/0092-8674(81)90187-2. PMID 6269758. Retrieved 2010-08-09.
- Alexeyev, Mikhail F. ; LeDoux, Susan P. ; Wilson, Glenn L. (July 2004). «Mitochondrial DNA and aging». Clinical Science 107 (4): 355–364. doi:10.1042/CS20040148. PMID 15279618.
- Reguly B, Jakupciak JP, Parr RL. 3.4 kb mitochondrial genome deletion serves as a surrogate predictive biomarker for prostate cancer in histopathologically benign biopsy cores. Can Urol Assoc J. 2010 Oct;4(5):E118-22. PMID PMID 20944788; PMID Central. PMID 2950771.
- Robinson K, Creed J, Reguly B, Powell C, Wittock R, Klein D, Maggrah A, Klotz L, Parr RL, Dakubo GD. Accurate prediction of repeat prostate biopsy outcomes by a mitochondrial DNA deletion assay. Prostate Cancer Prostatic Dis. 2010 Jun;13(2):126-31. Epub 2010 Jan 19. PMID. PMID 20084081.
- «نسخه آرشیو شده». بایگانیشده از اصلی در ۱۳ اوت ۲۰۱۱. دریافتشده در ۵ ژوئن ۲۰۱۱.
- Brown WM, George M Jr. , Wilson AC (1979). «Rapid evolution of mitochondrial DNA». Proc Natl Acad Sci USA 76 (4): 1967–1971. doi:10.1073/pnas.76.4.1967. PMC 383514. PMID 109836.
- Brown WM (1980). «Polymorphism in mitochondrial DNA of humans as revealed by restriction endonuclease analysis». Proc Natl Acad Sci USA 77 (6): 3605–3609. doi:10.1073/pnas.77.6.3605. PMC 349666. PMID 6251473.
- Paleo-DNA Laboratory - Forensic Services
- Gill P, Ivanov PL, Kimpton C, et al. (February 1994). «Identification of the remains of the Romanov family by DNA analysis». Nat. Genet. 6 (2): 130–5. doi:10.1038/ng0294-130. PMID 8162066.
- The details of the tests were published at Gil et al. , 'Identification of the Remains' The Duke of Fife was officially named as the source of the comparison sample of mtDNA in Ivanov, 'Mitochondrial DNA', p. 419.
انواع نوکلئیک اسیدها | |||||||
|---|---|---|---|---|---|---|---|
| مؤلفات سازنده | |||||||
| RNA (RNA کدکننده و غیرکدکننده) |
| ||||||
| DNA |
| ||||||
| آنالوگ |
| ||||||
| حاملان کلونینگ | |||||||