میتوکندری
میتوکندری (به انگلیسی: Mitochondrion) یا راکیزه[1] در یاخته (سلول)، اندامکی است که وظیفه آن تنفس سلولی و نوعی اندامک انتقال انرژی است که موجب میشوند انرژی شیمیایی موجود در مواد غذایی با عمل فسفوریلاسیون اکسیداتیو، به صورت پیوندهای پرانرژی فسفات ATP (آدنوزین تریفسفات) ذخیره شود. این اندامک در تمام یاختههای دارای تنفس هوازی به جز در باکتریها که آنزیمهای تنفسی آنها در غشای سیتوپلاسمی جایگزین شدهاند وجود دارد. میتوکندری نیز همانند کلروپلاست (سبزدیسه) از دو غشای داخلی و خارجی تشکیل شدهاست. با این تفاوت که دو غشای داخلی و خارجی فضای درون میتوکندری را به دو بخش تقسیم میکنند که عبارتند از: ۱- فضای درون میتوکندری ۲- فضای بین دو غشا. درون میتوکندری دی. اِن. اِی حلقوی نیز وجود دارد و میتواند مانند پلازمید باکتری بهطور مستقل از سلول نیز همانندسازی کند. درون میتوکندری مایعی سیّال به نام ماتریکس وجود دارد که واکنشهای مربوط به فرایند تنفس سلولی در آن انجام میشود. تنفس سلولی فرایندی است که طی آن انرژی ذخیره شده در غذاها (قندها) به اِی. تی. پی (مولکول سوختی سلول) تبدیل میشود. در غشای داخلی چینخوردگیهایی وجود دارد که به آن کریستا گویند قرار دارد که باعث افزایش سطح غشا میشود. همچنین ماده زمینه ای به نام ماتریکس که پر از مایع است پر شدهاست.
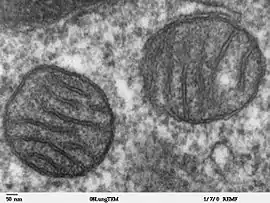
نام «میتوکُندری» ترکیبی است از دو واژه یونانی «میتوس»، μίτος، به معنای رشته و «خُندریون»، χονδρίον، به معنی دانه. چون این اندامک اغلب رشتهای یا به صورت دانههای کوچک در سیتوپلاسم همه سلولهای یوکاریوتی وجود دارد. میتوکندریها محل اکسیداسیون سلولی هستند. در طی این عمل، کربندیاکسید تولید میگردد و این همان کربندیاکسیدی است که از سلول دفع میشود. میتوکندری مانند کارخانهای باعث تولید انرژی سلول میشود. میتوکندری مربوط به بافت یونی پستانداران که ماتریس و غشای آنها توسط میکروسکوپ الکترونی نمایش داده شدهاست. اندامکی متصل به غشا است که در اکثر سلولهای یوکاریوتی (سلولهای یوکاریوتی سلولهایی هستند که دی. اِن. اِی آنها در هسته سلول قرار دارد و در اکثر گیاهان، حیوانات، قارچها و دیگر اشکال جانداران تشکیل میشوند) وجود دارند. کلمهٔ میتوکندری از عبارت یونانی Motos به معنای رشته …. به معنای دانه تشکیل شدهاست. اندازه میتوکندریها بین ۰٫۵ تا ۱٫۰ میکرومتر است. ساختارهای میتوکندری به عنوان نیروگاههای سلولی توصیف میشوند چرا که بیشتر انرژی شیمیایی سلول یا همان ATP از میتوکندری تولید میشود. علاوه بر تولید انرژی سلولی، میتوکندری وظایف دیگر درون سلولی نیز دارد که شامل سیگنالدهی، تمایز سلولی، مرگ سلولی و کنترل رشد و حفظ سلول هستند. میتوکندری میتواند در چندین نوع بیماری تأثیر داشته باشد که از جمله میتوان به اختلالات میتوکندری و اختلال عملکرد قلبی اشاره کرد. میتوکندری همچنین میتواند در پروسه پیری نیز مؤثر باشد. تحقیقات و پژوهشهای اخیر نشان میدهند که اوتیسم به ویژه اوتیسم حاد با نقص در سیستم میتوکندری در ارتباط است. شاخصههای متعدد و متفاوت، میتوکندریها را منحصر به فرد کردهاست. تعداد میتوکندریها در سلول بسته به نوع ارگانیسمها، بافتها و نوع سلول متفاوت است. برای مثال سلولهای خونی قرمز، فاقد میتوکندری هستند پس انرژی اش را از مسیر گلیکولیز به دست میآورد در حالی که سلولهای سازنده کبد بیش از ۲۰۰۰ میتوکندری در ساختار خود دارند. این اندامک شامل چندین توابع مختلف است که هر کدام عملکرد منحصر به فردی دارند. این واحد سلولی، شامل غشای خارجی، فضای بین دو غشا، کریستا و ماتریکس میتوکندری است. پروتئینهای میتوکندری بسته به نوع و بافت هر قسمت متفاوت هستند. در انسان، ۶۱۵ نوع مختلف از پروتئینهای میتوکندری قلبی شناسایی شدهاست. در حالی که در موشها این تعداد ۹۴۰ عدد است. پروتئومهای میتوکندری به شکل دینامیکی تنظیم میگردند اگرچه اکثر DNA موجود تصور میشود. در سلول دو نوکلئوس قرار دارد، اما میتوکندریها دارای ژنومهای مستقل خود هستند. علاوه بر این، DNA میتوکندری نشان میدهد که این واحد سلولی دارای شباهتهای قابل توجهی با ژنومهای باکتریایی است.
واژهشناسی
ساختواژه آن بهصورت [اسم(راک) + پسوند( ـ ایزه)] است. در ساخت این اسم از فرایند واژهسازی اشتقاق استفاده شدهاست. «راکیزه» از دو بخش «راک» به معنی رشته و نخ (در برابر «میتو» یونانی به همین معنی) و پسوند تصغیر «ـ ایزه» ساخته شدهاست. «ـ ایزه» دیس دیگر پسوندهای چه، ژه، ژک است که در واژگان بازارچه، نایژه، مژه، مژک بکار رفتهاند.
تاریخچه
اولین بررسیهای انجام شده بر روی میتوکندریها، در سال ۱۸۹۴ بهوسیله ریشارد آلتمان صورت گرفت که آنها را بیوپلاست یا جایگاههای زنده نامید؛ و نظر داد که بین واکنشهای اکسایش و کاهش سلول و میتوکندری وابستگی وجود دارد. در سال (۱۸۹۷) ابتدا با بررسیهای بیشتر آنها را میتوکندری نامید و در ۱۹۰۰، میکائیلیس به کمک معرف رنگی سبز ژانوس میتوکندری را در سلولهای زنده مشاهده کرد. واربورگ در سال ۱۹۱۳ آنزیمهای تنفسی را در این اندامک نشان داد. سرانجام برای اولین بار، در سال ۱۹۳۴، بنسلی و هر، توانستند آنها را از سلولهای کبدی جدا کرده و بعد آن بررسیهای بیشتر و عملیتر روی آن صورت گرفت. در سال ۱۹۱۳ ذرات گرفته شده از عصاره کبد خوکچه هندی نشان دادند که تنفس سلولی از طریق میتوکندری صورت میگیرد. واربورگ و هندیش در سال ۱۹۳۹ آزمایشهایی که با استفاده از ماهیچههای موش آزمایشگاهی انجام شدند، نشان دادند که یک اتم اکسیژن میتواند یک مولکول آذرونین تری فسفات تشکیل دهند. در سال ۱۹۴۱، اصطلاح پیوند فسفاتی نشان دهنده ایجاد انرژی در متابولیسم سلولی بود که توسط آلبرت لیمان ارائه شد. در سالهای بعد از آن مکانیسمی که در تنفس سلولی مورد بررسی قرار داده شد، توسعه یافت، هرچند ارتباط آن با میتوکندری هنوز هم مشخص نشده بود. در کتاب مقدمهای بر بخشهای سلولی نوشته آلبرت کلادیو، میتوکندری از لحاظ بیوشیمیایی و تحلیل از سایر بخشهای سلولی تفکیک شد. در سال ۱۹۴۶ اینطور نتیجهگیری شد که سیتوکروم اکسیداز و دیگر آنزیمها مسئولیت چرخه تنفسی سلولی را بر عهده دارند. اولین میکروگراف با وضوح بالا در سال ۱۹۵۲ تهیه شد و جایگزین زنجیره سبز ژانوس گردید که مطلوبترین شیوه مصورسازی میتوکندری بهشمار میرفت. این امر منجر به تحلیل جزئیات بیشتری از ساختار میتوکندری گردید که شامل غشای محصورکننده میتوکندری نیز است. همچنین مشخص گردید که غشای ثانویهای نیز در میتوکندری وجود دارد که بخشهای سختتر را از ساختار درونی جدا میسازد و در هر سلول دارای شکل خاصی است. اصطلاح معروف نیروگاه سلولی نیز توسط فیلیپ سیکوتیز در سال ۱۹۵۷ ابداع شد. در سال ۱۹۶۷ مشاهده شد که میتوکندریها حاوی ریبوزومها هستند. در سال ۱۹۶۸، روشهایی برای نقشهبرداری ژنهای موجود در میتوکندری توسعه یافتند و در سال ۱۹۷۶ با نقشههای ژنی، میتوکندری موجود در مخمرها تکمیل گردیدند.
شکل و اندازه میتوکندری و تغییرات آنها

شکل میتوکندریها متغیر اما اغلب رشتهای یا دانهای هستند. میتوکندریها در برخی مراحل عمل خود میتوانند به شکلهای دیگری درآیند. مثلاً، یک میتوکندری طویل ممکن است در یک انتهای خود متورم شده و به صورتی شبیه گرز درآید. (مثلاً در سلولهای کبدی چند ساعت بعد ورود غذا) یا ممکن است میان تهی شده و شکلی شبیه راکت تنیس به خود بگیرد. گاهی میتوکندریها حفره مانند شده و دارای بخش مرکزی روشنی میشود. اما بعد از مدتی، تمام این تغییرات به حالت اول برمیگردد.
اندازه
اندازه میتوکندریها نیز متغیر است و در بیشتر سلولها ضخامت آنها ۵۰µm و طول تا ۷µm میرسد. اما متناسب با شرایط محیطی و نیز مرحله عمل سلول، فرق خواهد کرد. سلولهایی که هم نوع هستند یا دارای عمل مشترک هستند دارای اندازه ثابت هستند.
ساختمان میتوکندری
غشای خارجی حدود ۷۵–۶۰ آنگستروم ضخامت دارد و از نوع غشاهای زیستی با ساختمان سه لایهای است. این غشا صاف و فاقد چین خوردگی است و هیچ ریبوزومی به آن نچسبیده، گاهی توسط شبکهٔ آندوپلاسمی احاطه میشود اما هیچگاه پیوستگی بین این دو دیده نشدهاست.
اتاق خارجی زیر غشای خارجی، فضایی در حدود ۲۰۰–۱۰۰ آنگستروم وجود دارد که به آن اتاق خارجی گفته میشود؛ که شامل دو بخش است: فضای بین دو غشا و فضای درون تاجها یا کریستاها یا کرتها. اما در برخی جاها غشای داخلی و خارجی بهم چسبیده و اندازه این فضا تقریباً صفر میشود. در این مناطق در مجاورت دو غشا، تراکمی از ریبوزومهای سیتوپلاسمی دیده میشود. به خاطر همین در نظر گرفته شده که این مناطق، محل عبور پروتئینهای مورد نیاز از سیتوزول به میتوکندری هستند. در این اتاق، ترکیباتی مثل آب، نمکهای کانی و یونها، پروتئینها، قندها، و چربیها SO2، O2، ATP و ADP وجود دارند. مقدار آب، بر اندازه کریستاها و در نتیجه بر ساخت ATP تأثیرگذار است.
غشای داخلی ضخامتش مثل غشای خارجی است اما ترکیب شیمیای آن فرق میکند. دارای چینخوردگیهای فراوانی است که به چینها، تاج یا کریستا گفته میشود. این چینها برخلاف سلولهای گیاهی، در سلولهای جانوری منظم قرار گرفتهاند.
اتاق داخلی فضای درونی میتوکندری که بهوسیله غشای داخلی دربرگرفته شده، اتاق داخلی گویند؛ که از ماده زمینهای با ماتریکس دربر گرفته شدهاست که ترکیب و ویژگیهای کلی آن، شبیه سیتوزول است و دارای آنزیمهای خاص و ریبوزوم خاص خود (۷۰S شبیه سلولهای پروکاریوتی) است. تعداد DNA، بر حسب نوع و سن سلول فرق میکند و مثل پروکاریوتها، دارای سیتوزین و گوانین زیادی است در نتیجه در مقابل گرما مقاوم است.
ژنوم میتوکندری
بررسیها نشان میدهد که DNAسازی در میتوکندری صورت میگیرد. طبق این بررسی به وجود DNA در میتوکندری پی میبریم. علاوه بر همانندسازی RNA و DNA سازی، پروتئینسازی هم در میتوکندری صورت میگیرد. این فرایند توسط آنزیمها و ملکولهای خاص خود اندامک صورت میگیرد. DNA میتوکندری اغلب موجودات حلقوی است. جایگاه DNA در ماده زمینه میتوکندری و بعضی مواقع چسبیده به غشای داخلی میتوکندری است. ژنوم میتوکندری سلولهای اغلب جانوران از ۲۰–۱۵ هزار جفت نوکلئوتید تشکیل یافتهاست و ژنوم میتوکندری در پستانداران حدود ۱۰۵ برابر کوچکتر از ژنوم هستهای است.
محصولاتی که توسط DNA میتوکندری رمز میشوند شامل RNAهای ریبوزومی میتوکندری tRNAها و برخی از پروتئینهای مسیر تنفس است. بعضی از پروتئینهای میتوکندری نیز در هسته رمز میشوند و پس از ساخته شدن در سیتوزول وارد اندامک میشوند. مثال مفروض از صفتی که توسط ژنوم میتوکندری تعیین میشود، جهت پیچش صدف در حلزون است که از وراثت سیتوپلاسمی تبعیت میکند. در حقیقت این صفات توسط ژنوم میتوکندری که همراه میتوکندریهای موجود در سیتوپلاسم وارد سلول تخم میشوند، انتقال مییابد و توارث به صورت تک والدی در اکثر آنها است.
نقش زیستی میتوکندری
- تنفس هوازی سلولها
تمام مواد انرژیزا، ضمن تغییرات متابولیکی درون سیتوپلاسمی با واسطه ناقلین اختصاصی به بستره میتوکندری میرسد. گلوکز بعد از تبدیل به استیل کو آنزیم A طی گلیکولیز به میتوکندری وارد میشود تا در چرخه کربس استفاده شود و اسیدهای چرب بهوسیله کارنی تین به داخل میتوکندری حمل شده که اینها هم سرانجام به استیل کو آنزیم A تبدیل میشوند. اسیدهای آمینه بعد از ورود به بستره به استیل کو آنزیم A تبدیل میشوند.
با انجام هر چرخه کربس که با استفاده از یک استیل کوآنزیم A در بستره میتوکندری آغاز میشود، علاوه بر CO2 و H2O سه مولکول نیکوتین آمید آدنین دی نوکلئوتید و یک مولکول FADH2 و یک مولکول GTP تولید میشود. این ناقلین انرژی در زنجیره انتقال الکترون استفاده شده و موجب تولید ATP میشوند.
- سنتز اسیدهای چرب
یکی از راههای تولید اسید چرب، سیستم میتوکندریایی است که عکس اکسیداسیون یا تجزیه آنها است.
- دخالت میتوکندری در گوارش چربیها
در هنگام گرسنگی، میتوکندریها به طرف ذرات چربی حرکت کرده و روی ذرات چرب خم شده و آنزیمهای میتوکندریایی شروع به هضم چربی و آزادسازی انرژی میکنند.
- ذخیره و تجمع مواد در میتوکندریها
میتوکندریها میتوانند در اتاق داخلی خود مواد مختلف را انباشته کنند که این مواد عبارتنداز از: ترکیبات آهندار، چربیها، پروتئینها، کاتیونها و آب. در اثر ذخیره این مواد، میتوکندریها اغلب به حالت یک غشایی و شبیه باکتریهای کوچک دیده میشوند و به تدریج، کریستاها محو میشوند اما بعد از حذف این مواد، دوباره همه به حالت اول برمیگردد.
- محل میتوکندریها در سلول
اغلب در اطراف هسته دیده میشوند اما در شرایط مرضی در حواشی سیتوپلاسم ظاهر میشوند. این پراکنش، تحت تأثیر مقدار گلیکوژن و اسید چرب میتواند قرار بگیرد. در طول میتوز میتوکندریها در مجاورت دوک جمع میشوند و وقتی تقسیم پایان مییابد، در دو سلول دختر، پراکنش تقریباً یکسانی پیدا میکند. پراکنش میتوکندریها را میتوان بر حسب عمل آنها از نظر تأمین انرژی، مطرح کرد که میتوکندریها در داخل سلولها جابجا شده و خود را به جایی که نیاز به ATP بیشتر است میرسانند.
- تعداد میتوکندریها در سلول
تشخیص ارزش میتوکندریایی یک سلول دشوار است. اما اغلب بر حسب نوع سلول مرحله عمل سلول متفاوت است. در یک سلول معمولی کبد بیشترین تعداد و در حدود ۱۰۰۰ تا ۱۶۰۰ عدد وجود دارد که در اثر تحلیل رفتن سلول و نیز سرطانی شدن آن کاهش مییابد؛ و در مقابل، تعداد میتوکندری در بافت لنفی، خیلی کمتر است. در سلولهای گیاهی، کمتر از جانوری است چون بسیاری از اعمال میتوکندریها، بهوسیله کلروپلاست انجام میشود.
منشأ میتوکندری
دو نظریه بیان شدهاست: یکی اینکه میتوکندریها ممکن است از قالبهای سادهتری ساخته شوند (تشکیل Denovo) و دیگر اینکه میتوکندریهای جدید از تقسیم میتوکندریهای قبلی به وجود میآیند. به این صورت که تعداد آنها، در طول میتوز و نیز در اینترفاز افزایش یافته و بعد بین دو سلول دختر، پراکنش مییابند.
- خاستگاه پروکاریوتی میتوکندری
فرضیهای در این صدد مطرح شدهاست که: در گذشته بسیار دور، جو زمین فاقد اکسیژن بوده و جاندارانی که در آن زمان میزیستهاند بیهوازی بودند. با گذشت زمان و ضمن واکنشهای شیمیایی، جو زمین دارای اکسیژن شده و به تدریج جانداران آن زمان و به ویژه پروکاریوتها به علت ساختمان ساده خود، هوازی شدهاند؛ بنابراین بعضی از باکتریها توسط سلولهای یوکاریوتی بلعیده شدند و به دلیل وجود همزیستی بعضی از آنها به کلروپلاست یا میتوکندری تبدیل شدند. پس یعنی اجداد میتوکندری همان پروکاریوتها یا باکتریها بودهاست.
حوای میتوکندریایی
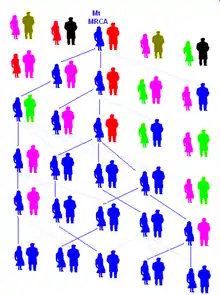
حوای میتوکندریایی به جدیدترین نیای مشترک تمام انسانهای امروزی از طرف مادری میگویند. دیانای میتوکندری که همواره از مادر به فرزند منتقل میشود، در همهٔ انسانهای امروزی بهطور مستقیم از حوای میتوکندریایی به ارث رسیدهاست.
حوای میتوکندریایی همتای مؤنث آدم کروموزوم Y (که جدیدترین نیای مشترک از سمت پدری است) است اما زمان زندگی این دو هزارها سال با هم متفاوت بودهاست. معمولاً تخمین زده میشود که حوای میتوکندری حدود ۲۰۰۰۰۰ سال پیش[2] و به احتمال زیاد در شرق آفریقا[3] زندگی میکردهاست. این همان زمانی است که گونه هومو ساپینس ساپینس (انسان دانای دانا یا همان انسان امروزی) در حال جدا شدن از دیگر گونههای انسانی بودهاست.
حوای میتوکندری خیلی زودتر از زمان هجرت از آفریقا که تصور میشود بین ۴۵٬۰۰۰ تا ۹۵٬۰۰۰ سال پیش رخ دادهاست، زندگی میکردهاست.[4] تعیین زمان زندگی 'حوا' ضربه بزرگی به این نظریه بود که نیاکان انسانهای امروزی که گونهای قدیمیتر از انسان مدرن بودند میلیونها سال پیش از آفریقا خارج شدهاند و در مناطق مختلف به نژادهای مختلف امروزی تکامل یافتهاند. این تخمین زمانی با نظریه مخالف منطبق است که میگوید انسان مدرن نسبتاً به تازگی در آفریقا سرچشمه گرفتهاست و از آنجا پراکنده شدهاست و جایگزین جمعیتهای انسانی «کهن» مانند نئاندرتالها شدهاست. فرضیه دوم در حال حاضر نظریه غالب است.
ترمیم چین و چروک پوست و ریزش مو با احیای عملکرد میتوکندری
با افزایش سن محتوای DNA میتوکندری کاهش مییابد به گونه ای که افراد به ازای هر ۱۰ سال، ۴ کپی از DNA میتوکندریایی خود را از دست میدهند. محققان به منظور بررسی تأثیر کاهش محتوای DNA میتوکندری در ظهور علائم پیری، یک جهش در دمین پلیمرازی آنزیم تکثیر کنندهٔ DNA میتوکندری ایجاد کردند. این جهش در تمامی جانوان باعث کاهش محتوای DNA میتوکندری میشود چرا که دقت در همانندسازی کاهش مییابد و جهشهای ایجاد شده طی همانندسازی، ۵۰۰ برابر بیشتر حفظ میشوند. سپس این ژن تغییر یافته را در پایین دست پروموتر تتراسایکلین قرار دادند و به یک تخم در مرحلهٔ تک سلولی وارد کردند. در ۸ هفتگی موشها به رژیم غذایی آنها تتراسایکلین اضافه کردند تا پروموتر فعال شود و رونویسی از ژن تغییر یافته صورت گیرد. با بیان ژن تغییر یافته در موشها، تغییراتی مانند چین و چروک پوست و ریزش مو ایجاد شد و موشهای جوان به موشهای پیر تبدیل شدند حتی باادامهٔ اضافه کردن تتراسایکلین و بیان ژن تغییر یافته، برخی از موشها مردند اما با حذف تتراسایکلین از رژیم غذایی موشها، به تدریج تغییرات ایجاد شده از بین رفتند و موشها دوباره جوان شدند. همچنین محتوای DNA میتوکندری که کاهش یافته بود مجدداً به حالت نرمال بازگشت. در نتیجه، برهم خوردن هومئوستازی DNA میتوکندری و در نتیجه اختلال در عملکرد میتوکندری مسئول ایجاد چین و چروک پوست و ریزش مو میباشد که با احیای عملکرد میتوکندری، تغییرات ایجاد شده بهبود مییابند.[5]
منابع
- واژهٔ مصوب فرهنگستان زبان و ادب فارسی، دفتر نخست تا چهارم، ۱۳۷۶ تا ۸۵
- دانشگاه لیدز -- تعیین زمان تاریخ مهاجرت انسان از طریق ساعت مولکولی
- سفری به ژنتیک -- پروژه Genographic
- Evaluating the mitochondrial timescale of human evolution", Trends Ecol. Evol. (Amst.) ۲۴ (۹), September ۲۰۰۹: ۵۱۵–۲۱
- Singh, B. , et al. , Reversing wrinkled skin and hair loss in mice by restoring mitochondrial function. Cell death & disease, 2018. 9(7): p. 735.
- کتاب زیستشناسی سال دوم تجربی.
- دانشنامه رشد.
| ||
| فرگشت |
|  |
| ژنتیک جمعیت | ||
| تکوین |
| |
| آرایه |
| |
| عضو |
| |
| فرآیند |
| |
| تمپوها و مدها |
| |
| گونهزایی |
| |
| تاریخچه اندیشه فرگشتی |
| |
| فلسفه |
| |
| مرتبط |
| |
| ||