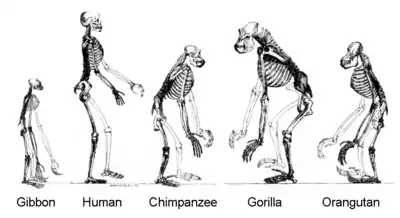
فرگشت انسان
با وجود این که شروع فرگشت انسان به عنوان یک موجود زنده، مانند تمام موجودات زندهٔ دیگر، به زمان پیدایش حیات بر روی کره زمین بازمیگردد؛ ولی بهطور کلی واژهٔ فرگشت انسان به تاریخچهٔ فرگشت نخستیسانان (پستانداران شبیه انسان) و به خصوص سرده انسان، از جمله پیدایش انسانها به عنوان گونهای مجزا از انسانسایان (کپیهای بزرگ) اطلاق میشود. مطالعهٔ فرگشت انسان زمینههای مختلف علمی از جمله انسانشناسی فیزیکی، نخستیشناسی، باستانشناسی، زبانشناسی، رویانشناسی، و ژنتیک را در بر میگیرد.[1]
| بخشی از مجموعه مقالههای |
| زیستشناسی فرگشتی |
|---|
 |
|
درگاه.ویکیپروژه |
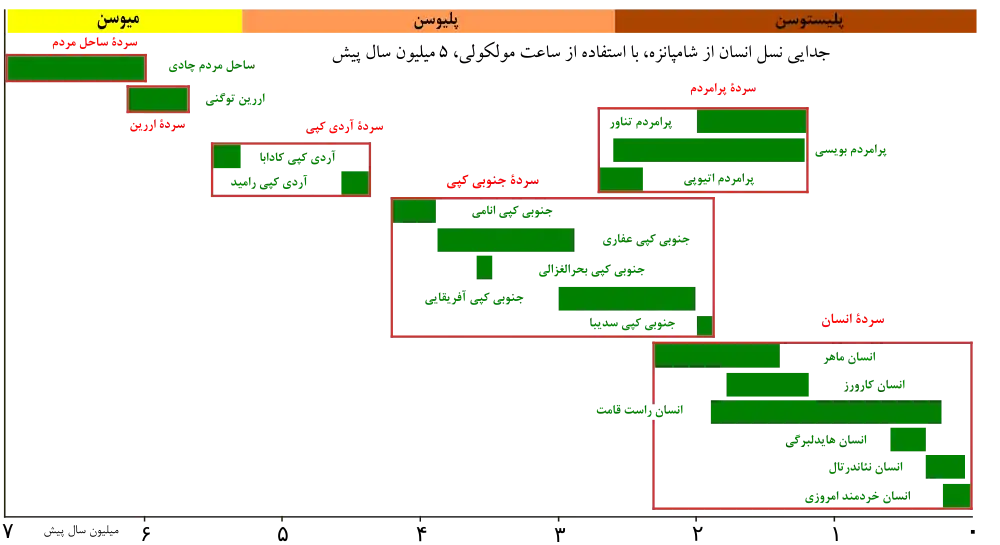
مطالعات ژنتیک نشان میدهند که فرگشت نخستیسانان احتمالاً در اواخر دورهٔ کرتاسه، ۸۵ میلیون سال پیش، شروع شدهاست. مجموعهٔ سنگوارههای گردآوری شده نیز بیانگر آنست که این زمان پیش از دورهٔ پالئوسن، ۵۵ میلیون سال پیش، بودهاست.[2][3] خانوادهٔ انسانسایان، یا کپیهای بزرگ، بین ۱۵ تا ۲۰ میلیون سال پیش از خانوادهٔ گیبونها جدا شدند. حدود ۱۴ میلیون سال پیش اورانگوتانها از خانوادهٔ انسانسایان جدا شدند.[4] راه رفتن روی دوپا ابتداییترین انطباق در دودمان انسانتباران است، و نخستین انسانتبارانی که بر روی دوپا راه میرفتهاند، ساحلمردم و اررین بودهاند. آردیکپی که کاملاً بر روی دوپا راه میرفتهاست، اندکی بعد فرگشت پیدا کردهاست. گوریلها و شامپانزهها حدوداً در یک زمان از این دودمان جدا شدند و احتمالاً ساحلمردم یا اررین آخرین نیای مشترک ما و آنها بودهاست. راه رفتن اولیه بر روی دو پا سرانجام منجر به فرگشت جنوبیکپیآسا و بعدها سرده انسان گردید. انسانهای امروزی از انسانسایانی که بین ۲٫۳ تا ۲٫۴ میلیون سال پیش در آفریقا میزیستند، تکامل یافتهاند.[5][6]
نخستین گونههای ثبت شده از سردهٔ انسان، انسان ماهر و انسان گاتنگی هستند که در حدود ۲٫۳ میلیون سال پیش تکامل پیدا کردند. بر طبق شواهد موجود، انسان ماهر اولین گونهای بوده که از ابزار استفاده میکردهاست. اندازهٔ مغز این انسانهای اولیه در حدود اندازهٔ مغز شامپانزه بودهاست. از آن زمان و در طول این چند میلیون سال تاکنون فرایند افزایش اندازهٔ مغز ادامه یافتهاست بهطوریکه با پیدایش انسان راستقامت، حجم جمجمهٔ آن به دو برابر حجم جمجمهٔ انسان ماهر افزایش یافته بود. انسان راستقامت و انسان کارورز نخستین انسانهایی بودند که به خارج از آفریقا کوچ کردند. این گونهها بین ۱٫۸ تا ۱٫۳ میلیون سال پیش در آفریقا، آسیا و اروپا پراکنده شدند. پنداشته میشود که این گونهها، نخستین گونههایی بودند که از آتش و ابزار پیچیده استفاده کردند.
دیدگاه غالب در میان دانشمندان این است که خاستگاه پیدایش انسان امروزی با فرضیه اخیر «خروج از آفریقا» یا «تک خاستگاهی» توضیح داده میشود[7][8][9] که بیان میدارد آغاز فرگشت انسانهای امروزی در آفریقا بودهاست و سپس بین ۵۰٫۰۰۰ تا ۱۰۰٫۰۰۰ سال پیش آنها به خارج از قارهٔ آفریقا مهاجرت کردند و کمکم جایگزین انسانهای راست قامت در آسیا و نئاندرتالها در اروپا شدند. فرضیهٔ دیگری نیز به نام «فرضیهٔ چند ناحیهای» وجود دارد که عنوان میکند انسانها پس از مهاجرت انسانهای راست قامت از آفریقا به سایر نقاط جهان در حدود ۲٫۵ میلیون سال پیش در مناطق جغرافیایی مختلف به صورت جمعیتهای جداگانه ولی با پیوند بین نژادها فرگشت یافتند. شواهد نشان میدهد که هاپلوتیپهای متعددی که منشأ نئاندرتالی دارند در حال حاضر در جمعیتهای غیرآفریقایی یافت میشود. شواهد نشان دهندهٔ آن است که نئاندرتالها و گونههای دیگر انسانسایان مانند انسانتبار دنیسووا تا ۶ درصد در ژنهای انسانهای امروزی (در نژادهای مختلف این مقدار متفاوت است) مشارکت داشتهاند.[10][11][12]
انسان امروزی که انسان خردمند نامیده میشود تنها گونهٔ بازمانده از انسانتباران است. انسانهای خردمند باستانی بین ۴۰۰٬۰۰۰ تا ۲۵۰٬۰۰۰ سال پیش فرگشت یافتند و از آنان انسانهای بهکالبد امروزی در اواسط دوره پارینه سنگی، حدود ۲۰۰٬۰۰۰ سال پیش تکامل پیدا کردند.[13] بنابر باور بسیاری از دانشمندان انسانهایی با رفتار نوین حدود ۵۰٬۰۰۰ سال پیش تکامل یافتند، هرچند که عدهای آغاز رفتار نوین در انسانها را همزمان با فرگشت انسانهای بهکالبد امروزی میدانند.[14]
تاریخچهٔ اندیشهها
آناکسیماندروس، فیلسوف یونان باستان عنوان کردهاست: «بشر در ابتدا به صورت حیوانی دیگر، شبیه ماهی بودهاست».[15]
کارل لینه کسی بود که برای اولین بار لغت homo را که امروزه در زیستشناسی (در مراجع انگلیسی) برای سردهٔ انسان به کار میرود را در سیستم طبقهبندی موجودات خود استفاده کرد. لینه و سایر دانشمندان هم عصرش کپیهای بزرگ را به عنوان نزدیکترین گونههای جانوری به انسان از نظر ریختشناسی و کالبدشناسی دستهبندی کردند. اندیشهٔ امکان وجود نیاکان مشترک برای انسان و کپیهای قدیم تر با انتشار کتاب خاستگاه گونهها توسط چارلز داروین در سال ۱۸۵۹ میلادی مطرح شد. داروین در این کتاب اندیشهٔ فرگشت گونههای جدید از گونههای قدیمتر را توضیح داد. کتاب داروین به منشأ پیدایش انسان پاسخی نداد و تنها به این نکته بسنده کرد که «به زودی منشأ پیدایش انسان و روند فرگشت آن روشن خواهد شد».

نخستین مناظرهها دربارهٔ ماهیت تکامل انسان بین توماس هاکسلی و ریچارد اون درگرفت. هاکسلی تکامل انسان از کپیها را با ارائه موارد بسیاری از شباهتها و تفاوتهای بین انسانها و کپیها توضیح داد. او همچنین در سال ۱۸۶۳ میلادی کتابی با عنوان شواهدی در مورد جایگاه انسان در طبیعت در این زمینه منتشر کرد. اما در هر حال خیلی از طرفداران اولیه داروین (مانند آلفرد راسل والاس و چارلز لایل) موافق نبودند که منشأ قابلیتهای ذهنی و احساسات اخلاقی انسانها میتواند با انتخاب طبیعی توضیح داده شود. داروین کاربرد نظریهٔ فرگشت و انتخاب جنسی در مورد انسانها را در کتابش با عنوان تبار انسان، و انتخاب طبیعی در ارتباط با جنسیت که در سال ۱۸۷۱ میلادی منتشر شد، توضیح داد.[16]
مشکل بزرگی که در آن زمان وجود داشت عدم وجود سنگوارههای گونههای انتقالی بود. با وجودی که اوژن دوبوا در سال ۱۸۹۱ میلادی سنگوارهٔ موجودی را در جزیرهٔ جاوه کشف کرد که هماکنون انسان راستقامت نامگذاری شدهاست، ولی این مشکل کماکان ادامه داشت تا اینکه در دههٔ ۱۹۲۰ میلادی با کشف نمونههای متعددی از سنگوارههای گونههای انتقالی در آفریقا، گردآوری اینگونه سنگوارهها آغاز شد. در سال ۱۹۲۵ میلادی، ریموند دارت گونه جنوبیکپی آفریقایی را از روی نمونه سنگوارهای مربوط به یک نوزاد که در اطراف شهر توانگ آفریقای جنوبی در یک غار پیدا شده بود، معرفی کرد. باقیماندههای این نوزاد یک جمجمهٔ بسیار کوچک و قالبی از مغز او بود. با وجودی که مغز یافت شده کوچک بود (۴۱۰ سانتیمتر مکعب)، ولی شکل آن برخلاف مغز شامپانزهها و گوریلها حالت گرد و به مغز انسانهای امروزی شباهت داشت. همچنین، نمونهٔ یافت شده دارای دندانهای نیش کوتاه بود و موقعیت سوراخ پس سری آن نشان میداد که این سنگواره مربوط به موجودی دو پا بودهاست. تمام این شواهد دارت را قانع ساخت که سنگوارهٔ کودک توانگ مربوط به نیاکان دو پای انسانها است که شکل انتقالی بین انسان و کپیها بودهاند.
طبقهبندی انسانها و وابستگان آنها از سال ۱۹۵۰ میلادی به بعد تغییرات عمدهای کردهاست.[17] به عنوان مثال، قبلاً جنوبیکپیآساهای ظریف به عنوان نیاکان سردهٔ انسان پنداشته میشدند[18] جنوبیکپیآساها و انسانهای خردمند امروزی هر دو از دودمان انسانتباران هستند.[19]
دادههایی که در دههٔ ۱۹۷۰ میلادی جمعآوری گردید نشان داد که جنوبیکپیآساها گروه متنوعی بودهاند و جنوبیکپیهای آفریقایی ممکن است نیاکان مستقیم انسان نبوده باشند.[20] با طبقهبندی مجدد جنوبیکپیآساها که در ابتدا به دو بخش ظریف و تنومند تقسیم میشدند، بخش تنومند به عنوان سردهای جدید با نام پرامردم معرفی شد.[20] آرایهشناسان، انسانهای خردمند امروزی، جنوبیکپیآساها و سایر گونههای وابسته را به مانند کپیهای بزرگ در خانواده انسانسایان قرار میدهند. ریچارد داوکینز در کتابش با نام داستان نیاکان عنوان میکند که جنوبیکپیآساهای تنومند نیاکان گوریلها بودهاند، پارهای از جنوبیکپیآساهای ظریف نیاکان شامپانزهها و باقی آنها نیاکان انسانهای خردمند امروزی بودهاند.[19]
پیشرفتهای عمده در دانش تعیین توالی دیانای در دهههای ۱۹۸۰ و ۱۹۹۰ میلادی، به ویژه در زمینهٔ دیانای میتوکندریایی (mtDNA) و دیانای Y یا Y-DNA کمک بزرگی به فهم منشأ پیدایش بشر کردهاست.[21][22][23] تعیین توالی mtDNA و Y-DNA که از گروههای گستردهای از جمعیتهای بومی گردآوری شد اطلاعات نیاکان آنها را که به ژنهای موروثی مردانه و زنانه مرتبط است، روشن ساخت.[24] مرتب کردن این اطلاعات در درخت ژنتیکی به عنوان پشتوانهای بر نظریهٔ خروج از آفریقا قلمداد شد.[25] تحلیلها نشان دهندهٔ گوناگونی بیشتر الگوی دیانای در آفریقا بودهاند که این مطلب با این نظریه که آفریقا موطن حوای میتوکندریایی و آدم کروموزوم Y بوده، سازگار است.[26]
شواهد موجود
شواهد فرگشت انسان در بسیاری از زمینههای علوم طبیعی یافت میشود. تا پیش از شروع قرن بیست و یکم بیشتر این شواهد در سنگوارههای گردآوری شده بود ولی با پیشرفتهای چشمگیر در زمینه ژنتیک در دو دهه اخیر، شواهد محکم ژنتیکی نیز در پشتیبانی از نظریه فرگشت بهطور عام و فرگشت انسان بهطور خاص به دست آمدهاست. آکادمی ملی علوم آمریکا در وب گاه خود عنوان میدارد که امروزه نظریه فرگشت به اندازه نظریه گردش زمین به دور خورشید توسط شواهد علمی پشتیبانی میشود و شواهد موجود به اندازهای است که دیگر این پرسش برای دانشمندان مطرح نیست که آیا انسان و سایر موجودات زنده محصول فرایند فرگشت هستند یا نه، بلکه پژوهشهای دانشمندان در مورد این است که جزئیات فرایند فرگشت چگونه بودهاست.[27] مطالعات خاص در مورد مبدأ حیات انسانها، انسانشناسی با گرایش دیرینانسانشناسی نامیده میشود.[28]
شواهد ژنتیکی
در ژنتیک تکاملی انسان بررسی میشود که تا چه میزان یک ژن انسان با بقیه و با گذشتهٔ فرگشتی که سبب پیدایش آن شده تفاوت دارد. تفاوت بین ژنها مفاهیم و کاربردهای متنوعی در انسانشناسی، پزشکی و پزشکی قانونی دارد. دادههای ژنتیک بینش عمدهای در زمینهٔ فرگشت انسان فراهم میکند. هر سال مقالات علمی متعددی در زمینه ژنتیک تکاملی انسان منتشر میشود. یکی از مهمترین این مقالهها در مجله علمی نیچر در سال ۲۰۰۵ به چاپ رسید. مقاله مذکور که نتایج پژوهش بیش از صد نفر از دانشمندان در ده مؤسسهٔ پژوهشی بزرگ دنیا بر روی ۲۳۷ میلیون جفت از اجزای تشکیلدهندهٔ کروموزوم شماره ۲ انسان است، نشان میدهد که این کروموزوم از جوش خوردن دو کروموزوم شامپانزه ایجاد شدهاست و به همین دلیل است که انسانها یک جفت کروموزوم کمتر از شامپانزهها دارند. نتایج این مقاله نشان میدهد انسان و شامپانزه نیاکان مشترک داشتهاند.[29]
شواهد فسیلشناسی
سنگوارههای گردآوری شده از انسانسایان نشان میدهد که در طول زمان به تدریج ستون فقرات آنان صاف شده، حجم مغز افزایش یافته، ترکیبات صورت ظریفتر شده، و عضلات مربوط به جویدن همزمان با تغییر وضعیت دندانها کمتر شدهاست. سنگوارههای بسیار زیادی از اجداد انسان در دورههای مختلف تاریخی جمعآوری شده که میتوان فهرستی از سنگوارههای مشهورتر را در صفحه فهرست سنگوارههای مشهور انسان در ویکیپدیا فارسی مشاهده نمود. همچنین میتوان فهرستی از سنگوارههای انسان همراه با توضیحات علمی مربوط به آن را در وب گاه موزه ملی تاریخ طبیعی (آمریکا) مشاهده نمود.[30]
دیگر شواهد
علاوه بر شواهد ژنتیکی و فسیل شناسی، شواهد دیگری نیز در آناتومی بدن انسان وجود دارد که نشان دهنده فرگشت انسان در طول تاریخ است.
در نخستیسانان عالی، دُم در لگن خاصره به صورت استخوان خاجی گنجانده شدهاست. تمام مهره داران حداقل در مدتی از زندگی خود دارای دُم هستند. در انسانهای خردمند امروزی، دُم به مدت چهار هفته در طول مراحل کارنگی هفتههای ۱۴ تا ۲۲ دورهٔ جنینی وجود دارد.[31] انسانهای خردمند امروزی دارای پلک سوم میباشند که کارایی ندارد. انسانها همچنین دارای عضلات خارجی گوش هستند[32] که حیوانات از آنها برای چرخاندن و کنترل گوشها به صورت مستقل از سر استفاده میکنند تا بتوانند بر صداهای خاصی متمرکز شوند. انسانها هنوز باقیماندههایی از چنین عضلاتی را دارند ولی این عضلات در طول روند فرگشت ضعیف شدهاند به گونهای که فقط قادرند گوشهای انسان را کمی تکان دهند.[33]
ماهیچه کفپایی در انسان نیز شاهدی است بر فرگشت انسانهای خردمند امروزی. این ماهیچه در حیوانات برای گرفتن و کنترل اشیاء با پاهایشان استفاده میشود و به آنها این قابلیت را میدهد که از پاهایشان مانند دستهایشان برای گرفتن اشیا استفاده کنند. در انسانها نیز این عضله وجود دارد ولی در فرایند فرگشت چنان کارایی خود را از دست دادهاست که امروزه پزشکان هنگام نیاز به بافتهای ترمیمی برای سایر قسمتهای بدن آن را خارج و از آن استفاده میکنند. این عضله چنان برای بدن انسان بیاهمیت شده که ۹٪ انسانهای امروزی بدون آن متولد میشوند.[34] مدرک دیگری بر فرگشت اندام ژاکوبسون است که بخشی از کالبد حیوانات میباشد. ریشهٔ این اندام را در انسان میتوان در پیشینهٔ جنسی انسانها در ماقبل تاریخ جستجو کرد. این اندام که در مجرای بینی واقع است، فرومونها را که مواد شیمیایی تحریککنندهٔ جنسی، هشدار دهنده یا یابندهٔ غذا هستند، ردیابی میکند. این اندام به حیوانات اجازه میدهد که همنوعانشان را برای جفتگیری یا اعلام خطر تعقیب کنند. انسانها با اندام ژاکوبسون به دنیا میآیند، ولی قابلیتهای این اندام در مراحل اولیهٔ پس از تولد رفته رفته کم میشود تا جایی که دیگر استفادهای ندارد.[35] در مواردی برای اندامهایی که به عنوان اندامهای زائد شناخته میشدند کاربردهایی پیدا شدهاست.[36] دندانهای عقل دلایل دیگری بر فرگشت انسان هستند. نیاکان انسان به مقدار زیادی از گیاهان تغذیه میکردند و نیاز داشتند تا مقادیر زیادی از گیاهان را در طول یک روز با سرعتی کافی بخورند تا بتوانند مواد مورد نیاز بدنشان را تأمین کنند. این موضوع خصوصاً زمانی اهمیت داشت که بدن انسان قابلیت هضم سلولز به میزان کافی را نداشت. هنگامی که عادات غذایی بشر تغییر کرد، آروارهٔ انسان کوچکتر شد و دیگر نیازی به دندان آسیای سوم یا دندان عقل نبود.[37]
در آوریل ۲۰۱۵ (اردیبهشت ۱۳۹۴) نتایج پژوهشی انتشار یافت که بیرونزدگی دیسک بینمهرهای که میتواند باعث کمردردهای مزمن شود در افرادی بیشتر دیده میشود که شکل مهرههای کمر آنها به مهرههای شمپانزهها و اورانگوتانها شباهت بیشتری دارد؛ به عبارت دیگر مهرههای کمر این افراد از نگاه فرگشتی کمتر برای راه رفتن روی دو پا تطبیق پیدا کردهاست. یکی از توضیحاتی که در این باره ارائه شده این است که در مسیر فرگشت از موجودی چهار دست و پا به انسان دو پا، فشار بر روی ستون مهرهها افزایش یافتهاست. نتایج این پژوهش این فرضیه را که کمردرد ممکن است منشأ فرگشتی داشته باشد تقویت میکند. شیوع بیماریهای ستون فقرات در انسان بیشتر از نخستیها است.[38][یادداشت 1]
پیش از انسان
فرگشت کپیهای بزرگ
تاریخچهٔ فرگشت نخستیسانان به ۶۵ میلیون سال پیش بازمیگردد.[39] قدیمیترین گونهٔ پستاندار شبیه به نخستیسانان،[40] آداپیسنما، ابتدا در آمریکای شمالی میزیست و بعدها در شرایط آب و هوایی گرمسیری دورهٔ پالئوسن و ائوسن در اوراسیا و آفریقا پراکنده گردید. نخستیسانان اولیه با توجه به شواهد موجود[41] در اوراسیا رشد و نمو پیدا کردند و این دودمان که بعدها منجر به پیدایش کپیهای آفریقایی و انسانها گردید، از اروپا و آسیای غربی به سمت جنوب به طرف آفریقا مهاجرت کردند. آن بخشی از جمعیت آنان، که بیشتر سنگوارههای آنان در لایههای بالایی دورهٔ ایوسن و لایههای پایینی دورهٔ الیگوسن در نزدیکی شهر فیوم مصر یافت شدهاست، در واقع نیاکان تمام گونههای زنده لمور در ماداگاسکار، چشمگرد تنبل در آسیای جنوب شرقی، شبدوست در آفریقا، راستبینیان یا میمونهای بر قدیم، میمونان: میمونهای بر جدید، کپیهای بزرگ و انسانهای خردمند امروزی محسوب میشوند.
قدیمیترین میمون بر قدیم شناخته شده کامویاکپی است که قدمت سنگوارههای یافت شدهٔ آن در لایههای بالایی دورهٔ الیگوسن در نواحی شمال کنیا ۲۴ میلیون سال برآورد میشود.[42] گمان میرود که نیای این میمون بر قدیم، گونهای نزدیک به آجیپتوکپی، پراپلیوکپی و پاراکپی باشد که در حدود ۳۵ میلیون سال پیش در فیوم میزیستهاست.[43] در سال ۲۰۱۰ میلادی، با معرفی سعدان که تخمین زده شده حدود ۲۸ تا ۲۹ میلیون سال پیش میزیستهاست، به عنوان گونهای نزدیک به آخرین نیای مشترک راستبینیان، جای خالی موجود در دورهای ۱۱ میلیون ساله در سنگوارههای گردآوری شده پر شد.[44]
.JPG.webp)
تنوع انواع درخت زی میمونهای بر قدیم و کپیها در شرق آفریقا در اوایل دورهٔ میوسن، حدود ۲۲ میلیون سال پیش، نشانگر تاریخچهای طولانی از پراکنده شدن گونهها از زمانهای پیشتر از آن است. سنگوارههایی که از ۲۰ میلیون سال پیش به جا ماندهاند شامل تکههایی هستند که به ویکتوریاکپی، قدیمیترین میمونهای بر قدیم نسبت داده میشوند. از میان سردههایی از دودمان کپیها که گمان میرود تا ۱۳ میلیون سال پیش میزیستهاند میتوان به پروکنسول، رنگواکپی، دندروکپی، لیمنوکپی، ناچولاکپی، اکواتوریوس، نیانزاکپی، افروکپی، هلیوکپی، و کنیاکپی که همگی در شرق آفریقا میزیستهاند اشاره کرد. وجود گونههایی غیر از میمونان بر قدیم در اواسط دوره میوسن و در مناطقی دور از هم، مانند اوتاوی کپی که سنگوارهٔ آن در رسوبات غارهای نامیبیا پیدا شده و پیرول کپی و درایوکپی که در فرانسه، اسپانیا و اتریش یافت شدهاند حکایت از تنوع فراوان این گونهها در آفریقا و مناطق مدیترانه در آب و هوای نسبتاً گرم و نامتغیر اوایل و اوسط دورهٔ میوسن دارد. جدیدترین گونه از خانوادهٔ انسانسایان در دورهٔ میوسن، اوپروکپی نام دارد که ۹ میلیون سال پیش میزیستهاست و سنگوارهٔ آن در بسترهای زغال سنگ در ایتالیا یافت شدهاست.
شواهد مولکولی نشان میدهد که گیبونها حدوداً ۱۲ تا ۱۸ میلیون سال پیش از کپیهای بزرگ جدا شدند و اورانگوتانها ۱۲ میلیون سال پیش از کپیهای بزرگ جدا شدند. هنوز سنگوارهای که بتوان با کمک آن نیاکان گیبونها را (که ممکن است محل پیدایش آنها از جمعیتهای انسانسایان جنوب شرق آسیا بوده باشد) مستند کرد، یافت نشدهاست. اما در مورد اورانگوتانها، میتوان سیواکپی در هند و گریفوکپی در ترکیه را که ۱۰ میلیون سال پیش میزیستهاند به عنوان نیاکان آنها برشمرد.[45]
جدا شدن دودمان انسان از سایر کپیهای بزرگ
از گونههای ناکالی کپی که سنگوارههای آن در کنیا و اورانوکپی که سنگوارههای آن در یونان یافت شدهاست، میتوان به عنوان گونههای نزدیک به آخرین نیای مشترک گوریلها، شامپانزهها و انسانها نام برد. شواهد مولکولی نشان میدهد که بین ۴ تا ۸ میلیون سال پیش، ابتدا گوریلها و سپس شامپانزهها از دودمانی که در نهایت به انسانهای خردمند امروزی انجامید، جدا شدند. از نظر مقایسهٔ چندریختی تک-نوکلئوتید تقریباً ۹۸٫۴ درصد دی ان ای انسان با دی ان ای شامپانزه یکسان است. سنگوارههای گردآوری شده از گوریلها و شامپانزهها محدود است. از جمله دلایل این موضوع، میتوان به شرایط نامطلوب جغرافیای محل زندگی آنان (خاک جنگلهای بارانی اسیدی است و استخوان را حل میکند) و خطای نمونهبرداری اشاره کرد.
سایر انسانساییان احتمالاً همراه با گونههای دیگر جانوران مانند بزکوهی، سگ، خوک، کفتار، فیل و اسب، به تدریج با آب و هوای خشکتر مناطق خارج از محدودهٔ نوار استوا تطبیق پیدا کردند. از حدود ۸ میلیون سال پیش، نوار استوایی (از نظر تقسیمبندی آب و هوایی) شروع به کوچکتر شدن کردهاست. سنگوارههای گونههای دودمان انسان پس از جدا شدن از شامپانزهها امروزه نسبتاً به خوبی شناخته شدهاند.
قدیمیترین آنان ساحلمردم چادی، که ۷ میلیون سال پیش و اررین توگنی که ۶ میلیون سال پیش میزیستند، هستند و پس از آنان سردههای زیر میزیستند:
- آردیکپی (۵٫۵ تا ۴٫۴ میلیون سال پیش) با گونههای آردیکپی کادابا و آردیکپی رامید
- جنوبیکپی (۴ تا ۱٫۸ میلیون سال پیش) با گونههای جنوبیکپی انامی، جنوبیکپی عفاری، جنوبیکپی آفریقایی، جنوبیکپی بحرالغزالی، جنوبیکپی گارهی، و جنوبیکپی سدیبا
- کنیامردم (۳ تا ۲٫۷ میلیون سال پیش) با گونهٔ کنیامردم پخترخ
- پرامردم (۳ تا ۱٫۲ میلیون سال پیش) با گونههای پرامردم اتیوپی، پرامردم بویسی، و پرامردم تناور
- انسانها (۲ میلیون سال پیش تاکنون) با گونههای انسان ماهر، انسان رودولفی، انسان کارورز، انسان گرجی، انسان پیشگام، انسان سپرانن سیس، انسان راستقامت، انسان هایدلبرگی، انسان رودزیایی، انسان نئاندِرتال، انسان خردمند ایدالتو، انسان خردمند باستانی، و انسان فلورسی
سردهٔ انسان

۱. گوریل ۲. جنوبی کپی ۳. انسان راستقامت ۴. نئاندرتال ۵.اشتاینهایم (مربوط به انسان خردمند باستانی یا انسان هایدلبرگی) ۶. انسان خردمند امروزی. کاهش پروگناتیزم و ضخامت خط ابرو، و افزایش اندازهٔ پیشانی قابل توجهاست.
انسان خردمند امروزی تنها گونهٔ بازمانده از سردهٔ انسان است. در حالی که برخی گونههای منقرض شدهٔ انسانها را میتوان به عنوان نیاکان انسانهای خردمند امروزی برشمرد، بسیاری دیگر احتمالاً تنها «خویشاوندان» ما بودهاند که از شاخههایی از نیاکان ما جدا شدهاند.[46][47] هنوز اجماعی بر اینکه کدامیک از این گروهها را باید به عنوان گونههای مجزا و کدامیک را به عنوان زیر گونه برشمرد، وجود ندارد. در برخی موارد این امر به دلیل کمیابی سنگوارهها و در برخی موارد به دلیل تفاوتهای جزئی در سیستمهای طبقهبندی گونهها برای سردهٔ انسان میباشد.[47] نظریه پمپ صحرای آفریقا یک توضیح محتمل برای گوناگونی اولیه سردهٔ انسان ارائه میدهد.
شواهد باستانشناسی و دیرینشناسی، امروزه این امکان را میدهد که تا حدودی در مورد رژیم غذایی گونههای متفاوت انسان استنباط نمود و نقش رژیم غذایی را در تکامل فیزیکی و رفتاری انسانها مطالعه کرد.[37][48][49][50][51]
انسان ماهر و انسان گاتنگی
انسان ماهر از ۲٫۴ تا ۱٫۴ میلیون سال پیش میزیستهاست. انسان ماهر در اواخر دورهٔ پالئوسن یا اوایل دورهٔ پلیستوسن، بین ۲٫۵ تا ۲ میلیون سال پیش، در جنوب و شرق آفریقا با جدا شدن از جنوبیکپیها تکامل پیدا کردهاست. انسان ماهر دندانهای آسیای کوچکتر و مغز بزرگتری از جنوبیکپیها داشته و ابزارهایی از سنگ و احتمالاً استخوانهای حیوانات میساختهاست. به دلیل استفاده از ابزار سنگی توسط این گونه که یکی از نخستین گونههای شناخته شدهٔ کپیهاست، کاشف آن، لوییس لیکی، نام مستعار «مرد ابزارکار» را برای آن برگزید. برخی از دانشمندان به دلیل اینکه اسکلت این گونه از نظر ریختشناسی به گونههای درختزی بیشتر شباهت دارد تا گونههای دوپایان، پیشنهاد کردهاند که این گونهها از سردهٔ انسان به سردهٔ جنوبی کپی منتقل شوند.[52]
انسان ماهر تا قبل از ماه میسال ۲۰۱۰ میلادی به عنوان اولین گونه از سردهٔ انسان پنداشته میشد، تا اینکه در این زمان سنگوارهٔ گونهای جدید از انسان به نام «انسان گاتنگی» در آفریقای جنوبی کشف شد که به احتمال فراوان پیش از انسانهای ماهر میزیستهاست.[53]
انسان رودولفی و انسان گرجی
اینها اسامی پیشنهاد شده برای گونههای مربوط به سنگواره انسانهایی هستند که حدود ۱٫۹ تا ۱٫۶ میلیون سال پیش میزیستهاند و ارتباطشان با انسان ماهر هنوز به درستی مشخص نیست.
- انسان رودولفی با یک جمجمه نیمه کامل یافت شده در کنیا شناسایی شدهاست. دانشمندان عنوان داشتهاند که این گونه نوعی انسان ماهر بودهاست، اما این موضوع هنوز ثابت نشدهاست.[54]
- انسان گرجی را که سنگوارهٔ آن در گرجستان پیدا شده، ممکن است بتوان به عنوان گونهای بینابینی بین انسان ماهر و انسان راستقامت،[55] یا زیرگونهای از انسان راستقامت قلمداد کرد.[56]
انسان کارورز و انسان راستقامت
اولین سنگوارههای انسان راستقامت توسط اوژن دوبوا، پزشک هلندی، در سال ۱۸۹۱ میلادی در جزیرهٔ جاوه کشور اندونزی پیدا شد. او ابتدا این گونه را بر اساس ویژگیهای ریختشناسی آن که حالتی بینابینی بین انسانها و کپیها داشت، انسان بوزینهٔ راست قامت نام نهاد.[57] انسان راستقامت بین ۱٫۷ میلیون تا ۷۰٫۰۰۰ سال سال پیش میزیستهاست (که میتوان نتیجهگیری کرد که در جریان فوران آتشفشان توبا از صفحهٔ روزگار محو شدهاست و از آن تنها گونههای انسان راستقامت سولویی و انسان راستقامت فلورسی به جا ماندند). اغلب از انسانهای مرحله اولیه این دوره به عنوان گونهای مجزا به نام انسان کارورز یا زیرگونهای از انسان راستقامت با عنوان انسان راستقامت کارورز یاد میشود.
گمان میرود که در اوایل دورهٔ پلیستوسن، ۱٫۵ تا ۱ میلیون سال پیش، جمعیتی از انسانهای ماهر با افزایش حجم مغز تکامل پیدا کردند و موفق به ساخت ابزارهای سنگی پیشرفته تری شدند؛ این تغییرات و پارهای دیگر از شواهد به گونهای است که انسانشناسان را متقاعد کردهاست که این گونهٔ تکامل یافته از انسانهای ماهر را به عنوان گونهای جدید به نام انسان راستقامت طبقهبندی کنند.[58] در این گونه زانوهای قفل شونده تکامل پیدا کردهاست و محل سوراخ بزرگ پسسری (سوراخی که طناب نخاعی از آن وارد جمجمه میشود) نیز عوض شدهاست. این گونه احتمالاً اولین گونهای بودهاست که گوشت را برای خوردن میپختهاست.
سنگوارهٔ مشهوری از انسانهای راستقامت، سنگوارهٔ انسان پکن است؛ نمونههای دیگری نیز در آسیا (به ویژه در اندونزی)، آفریقا، و اروپا یافت شدهاند. امروزه بسیاری از دیرینه شناسان از واژهٔ انسان کارورز برای بخشی از این گونه که در خارج از قارهٔ آسیا میزیستند استفاده میکنند و واژهٔ انسان راستقامت را برای آن بخشی که در آسیا میزیستهاند و مشخصات اسکلتی و دندانهای آنها اندکی با انسانهای کارورز تفاوت داشتهاست، به کار میبرند.
انسان سپرانن سیس و انسان پیشگام
این گونهها به عنوان گونههای بینابینی که در فاصله بین انسانهای راستقامت و انسانهای هایدلبرگی میزیستهاند، شناخته میشوند.
- انسان پیشگام از روی سنگوارههایی که در اسپانیا و انگلستان یافت شده، شناخته شدهاست و حیات آن به ۱٫۲ میلیون سال تا ۸۰۰٫۰۰۰ سال پیش بازمیگردد.[59][60]
- انسان سپرانن سیس از روی کاسهٔ جمجمهای که در ایتالیا پیدا شده و قدمت آن ۵۰۰٫۰۰۰ تخمین زده میشود، معرفی شدهاست.[61]
انسان هایدلبرگی
انسان هایدلبرگی (مرد هایدلبرگ) حدوداً بین ۶۰۰٫۰۰۰ تا ۳۵۰٫۰۰۰ سال پیش میزیستهاست. از این گونه به عنوان انسان خردمند هایدلبرگی نیز نام برده میشود.[62]
انسان رودزیایی و جمجمهٔ گویس
- تخمین زده میشود که انسان رودزیایی بین ۳۰۰٫۰۰۰ تا ۱۲۵٫۰۰۰ سال پیش میزیستهاست. بیشتر پژوهشگران امروز انسان رودزیایی را در گروه انسان هایدلبرگی طبقهبندی میکنند. با وجود این گروههای دیگری مانند انسان خردمند باستانی و انسان خردمند رودزیایی نیز برای این انسانها پیشنهاد شدهاست.
- در فوریه سال ۲۰۰۶ میلادی، جمجمهای به نام جمجمهٔ گویس کشف شد که احتمال میرود به گونهای بینابینی بین انسان راستقامت و انسان خردمند امروزی تعلق داشته باشد یا به یکی از گونههای متعددی که ادامه پیدا نکردهاند. قدمت این جمجمه که در اتیوپی کشف شدهاست بین ۵۰۰٫۰۰۰ تا ۲۵۰٫۰۰۰ سال برآورد میشود. در مورد این جمجمه جزئیات مختصری اعلام شدهاست و هنوز کاشفان آن در این مورد گزارش معتبر علمی منتشر نکردهاند.[63]
نئاندرتال و انسانتبار دنیسووا
انسان نئاندرتال که از آن با عنوان انسان خردمند نئاندرتال نیز نام برده میشود،[64] بین ۴۰۰٫۰۰۰[65] تا ۳۰٫۰۰۰ سال پیش در اروپا و آسیا میزیستهاست. شواهد مربوط به توالی دیانای میتوکندریایی نشان دهندهٔ آن است که هیچ شارش ژن عمدهای بین انسان نئاندرتال و انسان خردمند امروزی رخ ندادهاست که میتوان نتیجهگیری کرد که این دو، دو گونهٔ مجزا بودهاند که نیاکان مشترکی در حدود ۶۶۰٫۰۰۰ سال پیش داشتهاند.[66][67][68] به هر حال، توالی یابی ژن نئاندرتالها که در سال ۲۰۱۰ میلادی انجام شد، نشان داد که نئاندرتالها با انسانهای بهکالبد امروزی در حدود ۸۰٫۰۰۰ تا ۴۵٫۰۰۰ سال پیش (زمان تقریبی که انسانهای امروزی به خارج از آفریقا مهاجرت کردند ولی قبل از اینکه در اروپا، آسیا و جاهای دیگر پراکنده شوند)آمیزش داشتهاند.[69] بین ۱ تا ۴ درصد دیانای تقریباً تمام انسانهای غیر آفریقایی امروزی از دیانای نئاندرتال تشکیل شدهاست.[69] این دستاورد با مطالعات اخیر که نشان میدهد انشعاب برخی از اللهای انسان به یک میلیون سال پیش بازمیگردد سازگار است، هرچند که تفسیر نتایج این مطالعات مورد سؤال قرار گرفتهاست.[70][71] رقابت با انسانهای خردمند امروزی احتمالاً در انقراض نئاندرتالها نقش داشتهاست.[72][73] انسانهای خردمند امروزی و نئاندرتالها برای مدت ۱۰٫۰۰۰ سال در اروپا بهطور همزمان میزیستهاند.[74]

در سال ۲۰۰۸ میلادی، باستانشناسهایی که در غار دنیسووا در کوههای آلتای سیبری کار میکردند، بخشی از یک استخوان کوچک مربوط به انگشت پنجم یک نوجوان مربوط به گونهای از انسان را کشف کردند که اکنون با عنوان انسانتبار دنیسووا شناخته میشود.[75] وسایلی از جمله یک دستبند که در آن غار پیدا شده، مربوط به ۴۰٫۰۰۰ سال پیش میباشد. به دلیل هوای سرد آن غار، دیانای موجود در قطعه سنگواره سالم مانده بود در نتیجه هم دیانای میتوکندریایی و هم دیانای ژنوم هستهای در توالی سنجی بررسی شد.[10][76]
با وجودی که سرآغاز انشعاب دیانای میتوکندریایی انسانتبار دنیسووا بهطور غیرمنتظرهای به زمانهای قدیم بازمیگردد،[77] ولی توالی سنجی کامل ژنتیکی نشان دهندهٔ آن است که انسانتبار دنیسووا نیز به همان دودمان نئاندرتال تعلق دارد که زمان جدایی آن از گونهٔ نئاندرتال اندکی پس از این بوده که شاخهٔ نئاندرتال از شاخهٔ منتهی به انسانهای خردمند امروزی جدا شدهاست.[10] میدانیم که انسانهای امروزی در حدود ۱۰٫۰۰۰ سال در اروپا با نئاندرتالها در کنار هم میزیستهاند و کشف انسانتبار دنیسووا نیز این احتمال را مطرح کرد که این سه گونه یعنی انسان خردمند امروزی، نئاندرتالها و انسانتبار دنیسووا، در دورهای بهطور همزمان میزیستهاند. سوانت پابو، زیستشناس سوئدی، بیان کرده که وجود این زیر شاخه، تصویر زندگی انسانها را در اواخر دورهٔ پلیستوسن پیچیدهتر میکند.[75] همچنین شواهد نشان میدهد که امروزه حدود ۶ درصد از ژنهای مردم ملانزی از طرف انسانتبار دنیسووا میآید که این امر حکایت از آمیزش محدود انسانتبار دنیسووا با انسان خردمند امروزی در جنوب شرق آسیا دارد.[78]
اللهایی که گمان میرود از نئاندرتالها و انسانتبار دنیسووا سرچشمه گرفته باشد در محلهای مختلفی از ژنهای انسانهای خردمند امروزی خارج از قارهٔ آفریقا یافت شدهاست. انواع آنتی ژن لکوسیت انسان (HLA) که از طرف نئاندرتالها و انسانتبار دنیسووا آمدهاند بیش از نیمی از اللهای HLA مردم امروزی اوراسیا را تشکیل میدهند.[12]
انسان فلورسی
انسان فلورسی که بین ۱۰۰٫۰۰۰ تا ۱۲٫۰۰۰ سال پیش میزیستهاست، گاهی به طنز هابیت هم نامیده شدهاست که دلیل آن اندام کوچک این انسان به خاطر کوتولگی ناشی از انزوا میباشد.[79] انسان فلورسی یک گونهٔ شگفتانگیز هم از نظر اندازه و هم از نظر زمان زیست میباشد و از آن میتوان به عنوان نمونهای از گونههای اخیر سردهٔ انسان یاد کرد که خصوصیات تکامل یافتهٔ آن مشابه انسانهای امروزی نیست. به عبارت دیگر، انسان فلورسی دارای نیاکان مشترک با انسان خردمند امروزی بودهاست ولی پس از جدایی از شاخهٔ انسانهای امروزی، مسیر تکاملی جداگانهای را طی کردهاست. مهمترین اکتشاف در این مورد، سنگوارهٔ زنی ۳۰ سالهاست که در سال ۲۰۰۳ میلادی کشف شد و قدمت آن ۱۸٫۰۰۰ سال تعیین شدهاست. این زن، یک متر قد داشته و حجم جمجمهٔ آن تنها ۳۸۰ سانتیمتر مکعب بودهاست. (این حجم از حجم مغز شامپانزه نیز کوچکتر است و در حدود یک سوم حجم مغز انسانهای امروزی میباشد)
هم اکنون بحثهایی بین دانشمندان در جریان است راجع به این که آیا انسان فلورسی در واقع یک گونهٔ مجزا محسوب میشود یا نه.[80] برخی از دانشمندان بر این باورند که انسان فلورسی نوعی کوتوله (با دید آسیبشناسی) از انسان خردمند امروزی بودهاست.[81] این فرضیه تا حدودی دارای پشتوانهاست زیرا مردمی که هماکنون در فلورس، محلی که اسکلت انسان فلورسی در آن یافت شدهاست، زندگی میکنند دارای قد کوتاه هستند. مسئله دیگر در مورد انسان فلورسی نیز این است که سنگوارهٔ آن همراه با ابزارهایی که به انسانهای امروزی نسبت داده میشود، یافت شدهاست.[81]
فرضیه اینکه انسان فلورسی دچار کوتولگی از دید آسیبشناسی بودهاست، قادر به توضیح برخی ویژگیهای انسان فلورسی که نه تنها شبیه انسان امروزی نبوده بلکه بیشتر به نیاکان قدیمی سردهٔ انسان شبیهاست، نمیباشد. علاوه بر مشخصات جمجمه، این ویژگیها شامل شکل استخوانهای مچ، ساعد، بازو، زانو و پاها میگردد.
انسان خردمند امروزی
انسان خردمند امروزی از حدود ۲۵۰٫۰۰۰ هزار سال پیش تاکنون میزیستهاست. بین ۴۰۰٫۰۰۰ هزار سال پیش تا دومین دورهٔ بین یخبندانی در اواسط دورهٔ پلیستوسن، حدود ۲۵۰٫۰۰۰ هزار سال پیش، با پیدایش روند افزایش حجم جمجمه و پیشرفت در ساخت ابزار سنگی، زمینه برای فرگشت انسان خردمند امروزی از انسان راستقامت مهیا شد. شواهد مستقیم نشان میدهد که گروهی از انسانهای راستقامت ابتدا به خارج از آفریقا مهاجرت نمودند و باقی آنان که در آفریقا مانده بودند منجر به پیدایش انسان خردمند امروزی شدند. مهاجرتهای بعدی انسان خردمند امروزی در سطح قارهٔ آفریقا و به خارج از آن سرانجام باعث شد تا این گونه به تدریج جایگزین گونههای دیگر انسان راستقامت (مانند انسان نئاندرتال و انسانتبار دنیسووا) شود که زودتر به خارج از آفریقا کوچ کرده بودند. از این نظریه مهاجرت و پیدایش انسان امروزی غالباً با عنوان نظریهٔ «خروج از آفریقا» یاد میشود. به هر حال شواهد موجود نفیکنندهٔ فرگشت در نواحی مختلف یا برخیآمیزشهای انسان خردمند امروزی پس از مهاجرت به سرزمینهای جدید با جمعیتهای انسانی موجود در آن سرزمینها نیست. این موضوع هماکنون یک بحث داغ در علم دیرینمردمشناسی است.
پژوهشهای اخیر ثابت کردهاست که انسانها از نظر ژنتیکی بسیار یکدست هستند یا به عبارت دیگر دیانای افراد مختلف در انسان بیشتر از حد معمول در سایر گونهها به یکدیگر شبیهاست. این امر را میتوان ناشی از تکامل نسبتاً جدید در انسان یا ایجاد یک گلوگاه جمعیت حاصل از بلایای طبیعی همچون فوران آتشفشان توبا دانست.[82][83] به هر حال در انسانهای خردمند امروزی نیز ویژگیهای ژنتیکی مشخص در هر جمعیت وجود دارد که عمدتاً ناشی از مهاجرت گروههای کوچک از انسان خردمند امروزی در طول زمان به مناطق دارای شرایط آب و هوایی جدید میباشد. این خصوصیات تطابق یافته با محیط با اینکه بخش کوچکی از ژن انسان را تشکیل میدهد ولی شامل ویژگیهای مختلف بیرونی همچون رنگ پوست و شکل بینی و ویژگیهای درونی مانند قابلیت تنفس مؤثرتر در ارتفاعات، میگردد.
انسان خردمند ایدالتو زیرگونهای منقرض شده از انسان خردمند امروزی است که در حدود ۱۶۰٫۰۰۰ سال پیش در اتیوپی میزیستهاست.
- جدول مقایسهٔ گونههای انسان
| گونه | دوران زیست (میلیون سال پیش) | محل زندگی | قد فرد بالغ (متر) | وزن فرد بالغ (کیلوگرم) | ظرفیت جمجمه (۳ cm) | سنگوارههای موجود | تاریخ اکتشاف و تاریخ نامگذاری گونه (سال میلادی) |
|---|---|---|---|---|---|---|---|
| انسان گاتنگی | ۰٫۶–۲ | آفریقای جنوبی | ۱٫۰ | ۱ نمونه | ۲۰۱۰٫۲۰۱۰ | ||
| انسان ماهر | ۱٫۴–۲٫۳ | آفریقا | ۱–۱٫۵ | ۳۳–۵۵ | ۵۱۰–۶۶۰ | بسیار | ۱۹۶۴٫۱۹۶۰ |
| انسان کارورز | ۱٫۲۵–۱٫۸ | شرق و جنوب آفریقا | ۱٫۹ | ۷۰۰–۸۵۰ | بسیار | ۱۹۷۵ | |
| انسان رودولفی | ۱٫۹ | کنیا | ۱ جمجمه | ۱۹۸۵٫۱۹۷۲ | |||
| انسان راستقامت | ۰٫۲–۱٫۸ | آفریقا، اوراسیا (جاوه، هند، چین، قفقاز) | ۱/۸ | ۶۰ | ۸۵۰–۱۱۰۰ | بسیار | ۱۸۹۲٫۱۸۹۱ |
| انسان پیشگام | ۰٫۸–۱٫۲ | اسپانیا | ۱/۷۵ | ۹۰ | ۱۰۰۰ | ۲ نمونه | ۱۹۹۷ |
| انسان هایدلبرگی | ۰٫۳۵–۰٫۶ | اروپا، آفریقا، چین | ۱٫۸ | ۹۰ | ۱۱۰–۱۴۰۰ | بسیار | ۱۹۰۸ |
| انسان سپرانن سیس | ۰٫۳۵–۰٫۵ | ایتالیا | ۱۰۰۰ | ۱ کاسهٔ جمجمه | ۲۰۰۳٫۱۹۹۴ | ||
| انسان رودزیایی | ۰٫۱۲–۰٫۳ | زامبیا | ۱۳۰۰ | تعداد کمی | ۱۹۲۱ | ||
| انسان نئاندرتال | ۰٫۰۳–۰٫۳۵ | اروپا، غرب آسیا | ۱٫۶ | ۷۰–۵۵ | ۱۲۰۰–۱۹۰۰ | بسیار | ۱۸۶۴٫۱۸۲۹ |
| انسان خردمند امروزی | ۰٫۲ تاکنون | سراسر جهان | ۱٫۴–۱٫۹ | ۱۰۰–۵۰ | ۱۰۰۰–۱۸۵۰ | در حال زیست | - ۱۷۵۸٫ |
| انسان خردمند ایدالتو | ۰٫۱۵–۰٫۱۶ | اتیوپی | ۱۴۵۰ | ۳ جمجمه | ۲۰۰۳٫۱۹۹۷ | ||
| انسان فلورسی | ۰٫۰۱۲–۰٫۱۰ | اندونزی | ۱٫۰ | ۲۵ | ۴۰۰ | ۷ نمونه | ۲۰۰۴٫۲۰۰۳ |
| انسانتبار دنیسووا | ۰٫۰۴ | سرزمین آلتایی | ۱ نمونه | ۲۰۱۰ |
استفاده از ابزار
بهرهگیری از ابزارها، خود، نشانهای از هوشمندی است، پژوهشگران بر این باورند که کاربرد ابزار میتواند عامل محرک بخشهای ویژهای از تکامل انسان، به ویژه افزایش ادامه دار حجم مغز، تلقی گردد. با توجه به اینکه مغز انسان درصد قابل توجهی از انرژی بدن را مصرف میکند، نیاز انسانها در طول زمان و با افزایش حجم مغز به استفاده از غذاهای پرانرژی تر بیشتر شد. افزایش استفاده از ابزار به انسانها این امکان را داد که بتوانند فراوردههای گوشتی پرانرژی تری را از راه شکار به دست آورند و همچنین بتوانند گیاهان پرانرژی تر را پرورش دهند. پژوهشگران این مسئله را مطرح کردهاند که انسانسایان اولیه از نظر تکاملی تحت فشار بودهاند و مجبور به افزایش قابلیتهای خود برای ساخت و استفاده از ابزار شدهاند.[84]
اینکه انسانهای اولیه از چه زمانی شروع به استفاده از ابزار نمودهاند به سادگی قابل تعیین نیست. به دلیل اینکه هرچه این ابزارها ابتدایی تر باشند (مانند سنگهای دارای گوشههای تیز)، تشخیص دادن اینکه آنها طبیعی هستند یا ساختهٔ دست انسان، دشوارتر است. شواهدی در دست است مبنی بر اینکه ممکن است جنوبیکپیها (۴ میلیون سال پیش) از استخوانهای شکسته به عنوان ابزار استفاده میکردهاند ولی این موضوع مورد بحث است.[85]
باید توجه نمود که بسیاری از گونهها ابزار میسازند و از آن استفاده میکنند، ولی ساخت و استفاده از ابزارهای پیچیدهتر حیطهای در تسلط انسان هاست. قدیمیترین ابزارهای شناخته شده، «ابزارهای سنگی الدوایی» هستند که در اتیوپی پیدا شدهاند و مربوط به ۲٫۶ تا ۲٫۵ میلیون سال پیش میباشند که این زمان قبل از پیدایش نخستین گونههای سردهٔ انسان است. امروزه هیچ شواهدی در دست نیست که گونههایی از انسانها ۲٫۵ میلیون سال پیش میزیستهاند. سنگوارهای از انسان که در نزدیکی این ابزارها پیدا شدهاست و مربوط به ۲٫۳ میلیون سال پیش است، این احتمال را مطرح میکند که انسانها این ابزارها را ساختهاند. البته این موضوع در حد احتمال است و هنوز شواهد قوی در این مورد وجود ندارد. برنارد وود عنوان کردهاست که پرامردم تقریباً همزمان با گونههای اولیه انسان در دورهٔ ساخت ابزار الدوایی میزیستهاند. هرچند که مدرک مستقیمی مبنی بر استفادهٔ پرامردم از ابزار وجود ندارد، ولی خصوصیات بدنی آنان را میتوان به عنوان شاهد غیرمستقیمی برای قابلیت استفادهٔ آنان از ابزار در نظر گرفت. با این حال بیشتر دیرین انسانشناسان بر این باورند که گونههای اولیهٔ انسان بودهاند که ابزارهای الدوایی یافت شده را ساختهاند. آنها استناد میکنند که همواره سنگوارههای انسان، و نه پرامردم، در نزدیکی ابزارهای یافت شده پیدا شدهاند.[86]
در سال ۱۹۹۴ میلادی، رندال سوسمن با استناد به وضعیت شستها در دست، این بحث را مطرح کرد که هر دو سردهٔ انسان و پرامردم ابزار میساختهاند. او استخوانها و عضلات دست انسان را با شامپانزه مقایسه کرد و دریافت که انسانها دارای ۳ عضله در دست هستند که شامپانزهها از آن بیبهرهاند. همچنین دست انسان دارای قابلیتهای بهتری برای گرفتن اشیا نسبت به شامپانزهاست. سوسمن نشان داد که وضعیت شست دست در انسانهای امروزی پاسخی تکاملی به نیاز انسان برای ساخت و استفاده از ابزار بودهاست. او همچنین عنوان داشت که بر این اساس هر دو سردهٔ انسان و پرامردم ابزارساز بودهاند.[86]
- ابزار سنگی
با توجه به شواهد موجود، ابزار سنگی نخستین بار ۲٫۶ میلیون سال پیش توسط انسانهای ماهر استفاده گردید. این ابزار که به ابزارهای سنگی الدوایی موسوم هستند، از قسمت کردن سنگهای گرد توسط ضربه ساخته شدهاند.[87] ساختن این ابزار آغاز دورهٔ پارینهسنگی، یا عصر حجر قدیم تلقی میشود و پایان آن در آخر عصر یخبندان، حدود ۱۰٫۰۰۰ پیش در نظر گرفته میشود.
عصر پارینهسنگی به سه دورهٔ دیرینهسنگی (اوایل عصر پارینهسنگی که حدود ۳۵۰٫۰۰۰ تا ۳۰۰٫۰۰۰ سال پیش پایان مییابد)، میانسنگی (میانهٔ عصر پارینهسنگی که تا حدود ۵۰٫۰۰۰ تا ۳۰٫۰۰۰ سال پیش طول میکشد)، و نوسنگی تقسیم میشود.
دورهٔ بین ۷۰۰٫۰۰۰ تا ۳۰۰٫۰۰۰ سال پیش، به نام دورهٔ اشولی نیز نامیده میشود، زمانی که انسان کارورز تبرهای دستی بزرگی از جنس سنگ آتشزنه و کوارتزیت میساخت. این تبرها در اوایل دورهٔ اشولی ظاهری نخراشیده داشتند ولی بعدها به مرور زمان با پرداخت به وسیله ضربه زدن به لبههای تراشههای سنگی شکل ظریف تری به خود گرفتند. از ۳۵۰٫۰۰۰ هزار سال پیش، روش پرداخت دقیق تر سنگها شروع به توسعه یافتن کرد بهطوریکه با وارد کردن یک سری ضربات متوالی به سنگها، ابزارهایی برای خراش دادن، برش زدن و حتی سوزن ساخته شدند.[87] از ۵۰٫۰۰۰ سال پیش به این طرف، ابزارهای ظریف تر و تخصصی تری (مانند چاقو، تیغ و کفگیر) توسط نئاندرتالها و انسانهای مهاجر کرومانیون ساخته شد. در این دوره آنها همچنین شروع به ساخت ابزارهایی از استخوان نیز نمودند.



انسان خردمند امروزی و بحث «جهش بزرگ رو به جلو»

به نظر میرسد که روند استفاده از ابزار سنگی تا حدود ۴۰٬۰۰۰–۵۰٬۰۰۰ سال پیش به صورت پلکانی بودهاست. در هر مرحله (انسان ماهر، انسان کارورز، انسان نئاندرتال) استفاده از ابزار سنگی به صورت پیشرفته تری از مرحلهٔ قبل آغاز شدهاست، ولی بعد از شروع هر مرحله، توسعهٔ بیشتر ابزارها کند بودهاست. این گونههای انسان از لحاظ فرهنگی محافظه کار بودهاند، ولی از ۵۰٫۰۰۰ سال پیش، فرهنگ مدرن انسان شروع به رشد با سرعت بیشتری نمود. جارد دایموند، نویسندهٔ کتاب سومین شامپانزه، و سایر انسانشناسان این موضوع را «جهش بزرگ رو به جلو» مینامند.
انسانهای مدرن شروع کردند به دفن کردن مردگانشان، استفاده از پوست حیوانات برای پوشش، شکار با استفاده از روشهای ماهرانه تر (مانند استفاده از چاله به عنوان دام یا هدایت حیوانات به سمت پرتگاهها) و ایجاد غارنگارهها.[88] با پیشرفت فرهنگ انسانها، جمعیتهای مختلف انسانها شروع به نوآوری در فناوریهای موجود نمودند. در این زمان نوعی تنوع در ساخت مصنوعاتی مانند قلاب ماهیگیری، دکمه و سوزن استخوانی بین جمعیتهای مختلف انسان دیده میشود، چیزی که در جوامع انسانی قبل از ۵۰٫۰۰۰ سال پیش مشاهده نشدهاست. بهطور کلی، جوامع انسانهای نئاندرتال تنوع چندانی در تولید مصنوعاتشان نشان ندادهاند.
انسانشناسان مواردی چون تخصصی نمودن ابزارها، استفاده از زیورآلات و تصاویر (مانند نقاشی در غارها)، سازماندهی فضای زندگی، تشریفات (مانند خاک سپاری همراه با هدایا)، روشهای تخصصی شکار، کاوش مناطق جغرافیایی جدید، و ایجاد شبکههای معاملهٔ پایاپای را به عنوان نمونههای بارز از رفتار نوین انسانها میدانند. اینکه آیا پیدایش رفتار نوین در انسانها به صورت «انقلابی» بوده یا به تدریج اتفاق افتاده، مورد بحث است.[89]
مدلهایی در مورد پیدایش انسان

امروزه تمام انسانهایی که روی کرهٔ زمین زندگی میکنند به گونهٔ انسان خردمند امروزی تعلق دارند. اما بنا بر مدل «خروج از آفریقا»، این گونه اولین گونه از انسانها نبودهاست. نخستین گونه از سردهٔ انسان، انسان ماهر بودهاست که در حدود بیش از ۲ میلیون سال پیش در آفریقا تکامل یافتهاست. آنان در زمان نسبتاً کوتاهی در نواحی مختلف آفریقا پراکنده شدند. انسان راستقامت پیش از ۱٫۸ میلیون سال پیش تکامل پیدا کرد و تا ۱٫۵ میلیون سال پیش در تمام دنیای قدیم پراکنده شده بود.
برخی انسانشناسان معتقدند جوامع انسانی امروزی صرفاً در شرق آفریقا تکامل یافتند، گونهزایی کردند، و سپس به خارج از آفریقا کوچ نمودند و جایگزین جمعیتهای اوراسیا شدند (این فرضیه به مدل خروج از آفریقا یا «جایگزینی کامل» معروف است). برخی دیگر از انسانشناسان اعتقاد دارند انسانها به صورت یک جمعیت ولی در ارتباط تنگاتنگ با هم در نقاط مختلف تکامل پیدا کردند (این نظریه به فرضیه چند ناحیهای معروف است).
مدل خروج از آفریقا

بر پایهٔ مدل خروج از آفریقا، که توسط کریس استرینگر و پیتر اندروز ارائه شدهاست، انسان خردمند ۲۰۰٫۰۰۰ سال پیش در آفریقا تکامل پیدا کرد. انسانهای خردمند بین ۷۰٫۰۰۰ تا ۵۰٫۰۰۰ سال پیش شروع به مهاجرت به بیرون از قارهٔ آفریقا کردند و سرانجام جایگزین گونههای موجود انسانسایان در اوراسیا و آسیا شدند.[91][92] پژوهشهای انجام شده بر روی دیانای میتوکندریایی و کروموزوم Y مردانه فرضیهٔ خروج از آفریقا را تأیید میکنند. پس از تحلیل درختهای ژنتیکی که با استفاده از ۱۳۳ نوع از دیانای میتوکندریایی ساخته شدند، پژوهشگران به این نتیجه رسیدند که همهٔ آنها به یک نیای مادری مشترک در آفریقا بازمیگردد، که او را حوای میتوکندریایی نامیدند. این حقیقت که تنوع ژنتیکی میتوکندریایی در آفریقا بیشتر از سایر جاهاست نیز پشتوانهٔ دیگری برای فرضیهٔ خروج از آفریقا محسوب میشود.[93]
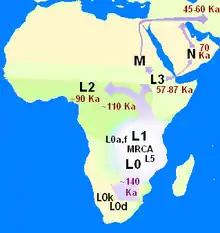
نظریههای مختلفی در مورد اینکه یک مهاجرت دسته جمعی به خارج از آفریقا صورت گرفتهاست یا چندین مورد، وجود دارد. یکی از مدلهایی که این پراکندگی را در چند مورد میداند، در برگیرندهٔ نظریهٔ پراکندگی جنوبی است،[95] که در سالهای اخیر توسط شواهد ژنتیکی، زبانشناسی و انسانشناسی نیز پشتیبانی شدهاست. در این نظریه، انسانهای نوین حدود ۷۰٫۰۰۰ سال پیش از راه سواحل شاخ آفریقا پراکنده شدند. این گروه به مسکونی شدن جنوب شرق آسیا و اقیانوسیه کمک کردند که این امر توضیح دهندهٔ اکتشاف مکانهای باستانشناسی مربوط با انسانهای اولیه در این مناطق بسیار پیشتر از اینگونه مکانها در سرزمین شام میباشد. موج دوم انسانها از راه شبهجزیره سینا به آسیا کوچ کردند که منجر به پیدایش جمعیت بزرگی از انسانها در اوراسیا گردیدند. این گروه دوم صنعت تولید ابزارهای پیچیده تری در اختیار داشتند و نسبت به گروه اول کمتر به منابع غذایی دریایی که آنان را محدود به زندگی ساحلی میکرد وابسته بودند. بیشتر شواهد مربوط به توسعهٔ گروه اول با بالا آمدن سطح آب دریا در پایان هر عصر یخبندان از بین رفتهاست.[95] مدل پراکندگی چند مرحلهای با مطالعاتی که نشان میدهد جمعیتهای اوراسیا و جمعیتهای جنوب شرقی آسیا و اقیانوسیه همگی از یک شاخهٔ دیانای میتوکندریایی هستند همخوانی ندارد.[96]
یک مطالعهٔ گسترده در مورد تنوع ژنتیکی در آفریقا، نشان داد که بوشمنها بیشترین تنوع ژنتیکی را در میان ۱۱۳ جمعیت مجزای مورد مطالعه دارا هستند. این موضوع نشان داد که آنان یکی از ۱۴ «خوشهٔ جمعیتی نیاکان» انسان هستند. در این پژوهش همچنین محل مبدأ مهاجرت انسانهای مدرن در جنوب غربی آفریقا، در نزدیکی مرز ساحلی نامیبیا و آنگولا تعیین شد.[97]
بر پایهٔ نظریهٔ فوران آتشفشان توبا که برخی از انسانشناسان و باستانشناسان آن را تأیید میکنند، فوران ابرآتشفشان دریاچهٔ توبا در جزیرهٔ سوماترا اندونزی در حدود ۷۰٫۰۰۰ سال پیش تبعات گستردهای در سطح جهان داشتهاست.[98] این فوران با کشتن بیشتر انسانهای زنده در آن زمان باعث ایجاد یک گلوگاه جمعیت شده و بر وراثت ژنتیکی تمام انسانهای حال حاضر کرهٔ زمین اثر گذاشتهاست.[99]
مدل چند ناحیهای

مدل چند ناحیهای، که مدلی است مبتنی بر الگوی تکامل انسان، در سال ۱۹۸۸ میلادی توسط میلفورد ولپاف ارائه شد.[100][101] تکامل چند ناحیهای بیان میدارد که تکامل انسان از ابتدای دورهٔ پلیستوسن، ۲٫۵ میلیون سال پیش، تاکنون در چهارچوب یک گونهٔ انسانی واحد، پیوسته در زمان و در سطح جهان از تکامل انسان راستقامت به انسان خردمند امروزی تکامل یافته، صورت گرفتهاست. بر اساس فرضیهٔ چند ناحیهای، دادههای ژنتیکی و سنگوارهای شواهدی بر تکامل انسان در سطح جهان هستند، و با گونهزایی اخیر که در فرضیهٔ خروج از آفریقا مطرح میشود، تناقض دارند. شواهد سنگوارهای برای ریچارد لیکی آنقدر کامل نبود که او بتواند این بحث را حل کند.[102] مطالعهٔ تکگروهها در دیانای میتوکندریایی و کروموزوم Y عمدتاً نظریهٔ خروج از آفریقا را تأیید کردهاند.[103] همچنین شواهد مربوط به دی ان ای اتوزومی غالباً موید فرضیه خروج ار آفریقاست؛ ولی به هر حال نشانههایی مبتنی برآمیزش باستانی در انسانهای امروزی در برخی مطالعات ارائه شدهاست.[104] توالییابیهای اخیر در مورد ژنهای نئاندرتال[105] و انسانتبار دنیسووا[106] نشان میدهد کهآمیزشهایی صورت گرفتهاست. انسانهای امروزی غیرآفریقایی ۴–۲ درصد از اللهای نئاندرتالها را در ژن خود دارند و برخی ملانزیها نیز %۶–۴ از اللهای انسانتبار دنیسووا را دارا هستند. این شواهد بانظریهٔ خروج ار آفریقا در تناقض نیستند مگر اینکه با دید خیلی سخت گیرانه تفسیر شوند. پس از احیا گلوگاه جمعیت ناشی از فوران ابرآتشفشان توبا، یک گروه نسبتاً کوچک از آفریقا به بیرون کوچ کردند و بهطور محدودی با نئاندرتالها، احتمالاً در خاورمیانه یا حتی پیش از خروج در شمال آفریقا،آمیزش داشتند و نوادگان عمدتاً آفریقایی آنان بودند که در سراسر جهان پراکنده شدند. یک بخشی نیز، احتمالاً در جنوب شرق آسیا و پیش از آنکه در ملانزی ساکن شوند با انسانتباران دنیسوواآمیزش داشتهاند.[107] هاپلوتیپهای HLA که منشأ نئاندرتال و انسانتبار دنیسووا دارند در جمعیتهای اوراسیایی و اقیانوسیهای امروزی یافت شدهاست.[12]
فرگشت اخیر و در حال رخداد در انسان
انتخاب طبیعی در جوامع انسانهای امروزی نیز رخ میدهد. موارد ثبت شدهای از تکامل در انسانهای امروزی شامل طولانی شدن دورهٔ بارداری و کاهش میزان کلسترول، قند خون و فشار خون میباشد.[108]
همچنین بحث تسریع تکامل انسان پس از (و در نتیجهٔ) توسعهٔ کشاورزی و تمدن در حدود ۱۰٫۰۰۰ سال پیش نیز مطرح است. ادعا میشود که این موضوع باعث ایجاد تفاوتهای اساسی ژنتیکی بین جوامع انسانی امروزی شدهاست.[109]
اولین انسان
از جمله پرسشهای اساسی که در طول تاریخ ذهن بشر را به خود مشغول داشته این است که اولین انسان چه کسی بودهاست؟ با پیشرفتهای حاصل شده در زمینهٔ زیستشناسی تکاملی، امروزه میتوان این مسئله را از نظر علمی مورد بررسی قرار داد. برای پاسخ به این پرسش باید توجه نمود که فرایند فرگشت در مورد همه موجودات و از جمله انسان فرایندی تدریجی، پیوسته و زمانبر است و در یک نسل اتفاق نمیافتد. ریچارد داوکینز این فرایند را به رشد انسان تشبیه میکند. همانگونه که شما در طول رشد نمیتوانید دقیقاً لحظهٔ تغییر از مرحلهٔ نوزادی به مرحلهٔ کودکی یا تغییر از کودکی به نوجوانی را تعیین کنید، در فرایند فرگشت نیز نمیتوانید دقیقاً یک موجود را از والدینش متمایز کرده، او را گونهای جدید بنامید. پیدایش گونهای جدید پدیدهای است که با گذشت زمانی طولانی و نه در یک نسل اتفاق میافتد؛ بنابراین پاسخ این پرسش را میتوان اینگونه بیان کرد: انسان خردمند امروزی نتیجه فرگشت میلیونها نسل از موجودات زنده از بدو پیدایش حیات است و در این میان هیچکدام از این موجودات را نمیتوان به عنوان اولین انسان محسوب کرد یا به عبارت دیگر اولین انسان هیچگاه وجود نداشتهاست. [110]
نظر ادیان
اسلام
به گفته سید محمدحسین طباطبایی دیدگاه دین اسلام دربارهٔ آفرینش انسان، با نظریه تکامل متفاوت است. براساس آیات قرآن، اولین انسانها آدم و حوا بودهاند[111] و انسانهای فعلی از نسل آنها هستند. اما در قرآن و روایات شواهدی وجود دارد که میگوید پیش از خلقت آدم و حواء، انسانهای دیگری نیز وجود داشتهاند. اما خلقت آنها با خلقت آدم و حواء جدا بودهاست و آدم و حواء از نسل انسانهای پیشین نیستند.[112] الله موجوداتی شبیه انسان خلق کرده بود که قبل از خلقت آدم و حواء از بین رفتند.
در تفسیر المیزان، علامه طباطبایی به نقل از محمد باقر (پنجمین امام شیعیان) آوردهاست: «خداوند عز و جل در همین زمین از روزی که آن را آفریده، هفت عالم خلق کرده (و سپس برچیده) و هیچیک از آن عوالم از نسل آدم ابوالبشر نبودند و خدای تعالی همهٔ آنها را از پوستهٔ روی زمین آفرید و نسلی را بعد از نسل دیگر ایجاد کرد و برای هر یک عالمی بعد از عالم دیگر پدیدآورد تا در آخر آدم ابوالبشر را بیافرید و ذریهاش را از او منشعب ساخت.»[113]
پژوهشگران برجسته در زمینهٔ فرگشت انسان
- چارلز داروین، طبیعیدان بریتانیایی که شواهد عمدهای را در مورد اینکه گونهها در اثر تغییرات تکاملی ایجاد میشوند، گردآوری کرد.
- کریس استرینگر، انسانشناس بریتانیایی، از پیشگامان مطرحکنندهٔ فرضیهٔ خروج از آفریقا
- رابرت بروم، پزشک و دیرینشناس اسکاتلندی که پژوهشهای او در آفریقای جنوبی منجر به کشف سردهٔ پرامردم و جمجمهٔ خانم پلس گردید.
- سوانت پابو، زیستشناس سوئدی متخصص در زمینهٔ ژنتیک تکاملی
- دیوید پیلبیم، دیرینمردمشناس، پژوهشگر و نویسندهٔ آمریکایی در زمینهٔ تکامل انسان و نخستیسانان
- آلن تمپلتون، پژوهشگر ژنتیک و آمار آمریکایی، از پیشگامان مطرحکنندهٔ فرضیهٔ چند ناحیهای
- فیلیپ توبیاس، دیرینمردمشناس آفریقای جنوبی از برجستهترین پژوهشگران تکامل انسان در جهان محسوب میشود.
- اریک ترینکاس، دیرینمردمشناس آمریکایی متخصص در زمینهٔ زیستشناسی نئاندرتالها و تکامل انسان
- دونالد جانسون، دیرینمردمشناس آمریکایی، کاشف گونهٔ جنوبیکپی عفاری
- ریموند دارت، کالبدشناس و دیرینمردمشناس استرالیایی که کارهای پژوهشی او در توانگ آفریقای جنوبی، به کشف گونهٔ جنوبیکپی آفریقایی انجامید.
- ریچارد داوکینز، زیستشناس تکاملی و نویسنده
- جفری شوارز، انسانشناس آمریکایی و استاد انسانشناسی زیستی
- استفان جی گولد، دیرینشناس، زیستشناس تکاملی و مورخ علم اهل آمریکا
- جفری لیتمن، کالبدشناس و انسانشناس آمریکایی که در مورد تکامل اصوات و صحبت کردن پژوهش نمودهاست.
- لوییس لیکی، انسانشناس و طبیعی دان کنیایی که در مبحث تکامل انسان در آفریقا فعالیت کردهاست.
- ماری لیکی، باستانشناس و انسانشناس بریتانیایی که از جمله اکتشافات او در آفریقا میتوان به ردپاهای لتولی اشاره کرد.
- ریچارد لیکی، دیرینمردمشناس و انسانشناس کنیایی، فرزند لوییس و ماری لیکی
- هنری مک هنری، انسانشناس آمریکایی که تخصص او در زمینهٔ تکامل انسان، پیدایش راه رفتن بر روی دو پا و دیرینمردمشناسی است.
- میلفورد ولپاف، دیرینمردمشناس آمریکایی، از پیشگامان مطرحکنندهٔ فرضیهٔ چند ناحیهای
فهرست گونهها
|
|
جستارهای وابسته
یادداشت
- در این پژوهش آنها رابطه بین شکل مهره، حرکت و جابجایی آن و زائده اشمورل (Schmorl’s node) را مورد مطالعه قرار دادند. زائده اشمورل بیرونزدگی غضروف (دیسک) بین مهرهها به داخل بافت استخوانی مهره بالایی یا پایینی است. این زائده معمولاً دردناک نیست، بیشتر در سنین بالا دیده میشود و محصول فرسودگی مهره در گذر عمر است. این مطالعه نشان داد در کسانی که شکل مهرهها بیشتر به شکل مهرههای اجداد ما شبیه است این فتق عمودی بین مهرهای بیشتر دیده میشود. علاوه بر این مهره اول کمر در صورت سلامت، به راحتی از مهره مشابه در شمپانزهها و اورانگوتانها قابل بازشناسی است اما اگر این مهره سالم نباشد این تمایز دیده نمیشود؛ به عبارت دیگر زائده اشمورل مهره انسانی را به مهره نخستیها شبیه میکند.
منابع
- Heng, HH (2009). "The genome-centric concept: resynthesis of evolutionary theory". Bioessays. ۳۱ (۵): ۵۱۲–۲۵. doi:10.1002/bies.200800182. PMID 19334004. Unknown parameter
|ماه=ignored (help);|access-date=requires|url=(help) - "Nova – Meet Your Ancestors". PBS. Retrieved 2008-10-24.
- "Plesiadapis" (PDF). North Dakota Geological Survey. Retrieved 2008-10-24.
- Dawkins R (2004) The Ancestor's Tale.
- "Query: Hominidae/Hylobatidae". Time Tree. 2009. Retrieved December 2010.
- Stringer, C.B. (1994), "Evolution of Early Humans", in Jones, Steve; Martin, Robert; Pilbeam, David, The Cambridge Encyclopedia of Human Evolution, Cambridge University Press, p. ۲۴۲, ISBN 978-0-521-32370-3
- McHenry, H.M (2009), "Human Evolution", in Ruse, Michael; Travis, Joseph, Evolution: The First Four Billion Years, Cambridge, Massachusetts: The Belknap Press of Harvard University Press, p. ۲۶۵, ISBN 978-0-674-03175-3
- "Out of Africa Revisited - 308 (5724): 921g - Science". Sciencemag.org. 2005-05-13. doi:10.1126/science.308.5724.921g. Archived from the original on 22 November 2010. Retrieved 2009-11-23.
- Nature (2003-06-12). "Access: Human evolution: Out of Ethiopia". Nature. Archived from the original on 22 November 2010. Retrieved 2009-11-23.
- "Origins of Modern Humans: Multiregional or Out of Africa?". ActionBioscience. Archived from the original on 22 November 2010. Retrieved 2009-11-23.
- Reich D, Green RE, Kircher M, et al. (2010). "Genetic history of an archaic hominin group from Denisova Cave in Siberia". Nature. ۴۶۸ (۷۳۲۷): ۱۰۵۳–۶۰. doi:10.1038/nature09710. PMID 21179161. Unknown parameter
|month=ignored (help) - Noonan J. P (2010). "Neanderthal genomics and the evolution of modern humans". Genome Res. ۲۰ (۵): ۵۴۷–۵۳. doi:10.1101/gr.076000.108. PMID 20439435. Unknown parameter
|month=ignored (help) - Laurent Abi-Rached, et al. (2011-08-25). "The Shaping of Modern Human Immune Systems by Multiregional Admixture with Archaic Humans". Science. ۳۳۴ (۶۰۵۲). doi:10.1126/science.1209202. PMID 21868630. Archived from the original on Aug 2011.
- Fossil Reanalysis Pushes Back Origin of Homo sapiens, February 17, 2005, Scientific American.
- Mellars, Paul (2006). "Why did modern human populations disperse from Africa ca. 60,000 years ago?". Proceedings of the National Academy of Sciences 103 (25): 9381–6. Bibcode 2006PNAS..103.9381M. doi:10.1073/pnas.0510792103. PMC 1480416. PMID 16772383.
- بنا بر نوشتهٔ هیپولیتوس رومی، کتاب یکم
- Darwin, Charles (1871), The Descent of Man, and Selection in Relation to Sex, Princeton NJ: Princeton University Press, p. ۲۴۲, ISBN 0-691-02369-7
- Walter, Carl Hartwig (2002), The primate fossil record, Cambridge University Press, p. ۴۰۹, ISBN 978-0-521-66315-1, retrieved 6 November 2011
- Grine, Frederick (31 December 2007), Evolutionary History of the "Robust" Australopithecines, Transaction Publishers, p. ۳۰۱, ISBN 978-0-202-36137-6
- Dawkins, Richard; Wong, Yan (September 2005), The Ancestor's Tale: A Pilgrimage to the Dawn of Evolution, Houghton Mifflin Harcourt, p. ۸۶, ISBN 978-0-618-61916-0
- Philip Briggs (1 October 2009), Tanzania: With Zanzibar, Pemba & Mafia, Bradt Travel Guides, p. ۶, ISBN 978-1-84162-288-0
- M'charek, Amade (2005), The Human Genome Diversity Project: an ethnography of scientific practice, Cambridge University Press, p. ۹۶, ISBN 978-0-521-83222-9
- DeSalle, Rob; Tattersall, Ian (2008), Human origins: what bones and genomes tell us about ourselves, Texas A&M University Press, p. ۱۴۶, ISBN 978-1-58544-567-7
- Trent, R. J. (2005), Molecular medicine: an introductory text, Academic Press, p. ۶–, ISBN 978-0-12-699057-7
- Webster, Donovan; Wells, Spencer (20 April 2010), Meeting the Family: One Man's Journey Through His Human Ancestry, National Geographic Books, p. ۵۳, ISBN 978-1-4262-0573-6
- Speicher, Michael R.; Motulsky, Stylianos E. (3 February 2010), Vogel and Motulsky's Human Genetics: Problems and Approaches, Springer, p. ۶۰۶, ISBN 978-3-540-37653-8
|first3=missing|last3=in Authors list (help) - Adam's Gene and the Mitochondrial Eve، Xlibris Corporation، ۱۴ سپتامبر ۲۰۰۹، ص. ۴۰، شابک ۹۷۸-۱-۴۴۱۵-۰۷۲۹-۷ پارامتر
|first1=بدون|last1=در Authors list وارد شدهاست (کمک) - human evolution
- Fossil Hominids: the evidence for human evolution
- L.W. Hillier et al. (2005),"Generation and annotation of the DNA sequences of human chromosomes 2 and 4", Nature, 434, 724-731
- Saraga-Babić, M; Lehtonen, E; Svajger, A; Wartiovaara, J (1994). "Morphological and immunohistochemical characteristics of axial structures in the transitory human tail". Annals of anatomy. ۱۷۶ (۳): ۲۷۷–۸۶. PMID 8059973.
- Owen, R. 1866–1868. Comparative Anatomy and Physiology of Vertebrates. London
- Montagna, W; Machida, H; Perkins, E.M. (1966). "The skin of primates XXXIII. : The skin of the angwantibo". American Journal of Physical Anthropology. ۲۵ (۳): ۲۷۷–۲۹۰. doi:10.1002/ajpa.1330250307. PMID 5971502.
- Wiedersheim, Robert (1893). The Structure of Man: an index to his past history. London: Macmillan and Co.
- Muller, G. B. (2002) "Vestigial Organs and Structures." in Encyclopedia of Evolution. Mark Pagel, editor in chief, New York: Oxford University Press. pp. 1131–1133
- "Vestigial Organs Not So Useless After All, Studies Find"
- Walker, Alan (2006). "Early Hominin Diets: Overview and Historical Perspectives". In Peter Ungar. Evolution of the Human Diet: The Known, the Unknown, and the Unknowable. US: Oxford University Press. pp. ۳–۱۰. ISBN 978-0-19-518346-7. (scroll up to view chapter 1 & part of chapter 2, which is a serendipitous result from another search. Subsequent attempts get a targeted search result gave returns without chapter 1).
- The ancestral shape hypothesis: an evolutionary explanation for the occurrence of intervertebral disc herniation in humans BMC Evolutionary Biology 2015
- page 9 ,cf.osb&fp=3640775752d61e11&biw=1600&bih=721 1999] - [P.R. Willoughby - 2005 International Journal of Comparative Psychology UCLA] - [R.D. Martin - 2002 doi:10.1016/B0-08-043076-7/03083-7 - [S. Tavaré, C. R. Marshall, O. Will, C. Soligo, & R. D. Martin - 2001 doi:10.1038/416726a [Retrieved 2012-01-01]
- [K. D. Rose -1994 DOI: 10.1002/evan.1360030505 - [J.Fleagle,C.Gilbert 2011-2012 - [J.Roach 2008 - [V. McMains - 2011 بایگانیشده در ۱۶ ژانویه ۲۰۱۹ توسط Wayback Machine - [2009 & [Retrieved 2012-01-01]
- Kordos L, Begun D R (2001). "Primates from Rudabánya: allocation of specimens to individuals, sex and age categories". J. Hum. Evol. ۴۰ (۱): ۱۷–۳۹. doi:10.1006/jhev.2000.0437. PMID 11139358.
- David W. Cameron (2004). Hominid adaptations and extinctions. UNSW Press. p. ۷۶. ISBN 978-0-86840-716-6. Retrieved 6 November 2011.
- David Rains Wallace (13 September 2005). Beasts of Eden: Walking Whales, Dawn Horses, and Other Enigmas of Mammal Evolution. University of California Press. pp. ۲۴۰–. ISBN 978-0-520-24684-3. Retrieved 6 November 2011.
- Zalmout, I.S.; Sanders, W.J.; MacLatchy, L.M.; Gunnell, G.F.; Al-Mufarreh, Y.A.; Ali, M.A.; Nasser, A. -A.H.; Al-Masari, A.M.; Al-Sobhi, S.A. (2010). "New Oligocene primate from Saudi Arabia and the divergence of apes and Old World Monkeys". Nature. ۴۶۶ (۷۳۰۴): ۳۶۰–۳۶۴. doi:10.1038/nature09094. PMID 20631798.
- Srivastava (2009). Morphology Of The Primates And Human Evolution. PHI Learning Pvt. Ltd. p. ۸۷. ISBN 978-81-203-3656-8. Retrieved 6 November 2011.
- Strait DS, Grine FE, Moniz MA (1997). "A reappraisal of early hominid phylogeny". J. Hum. Evol. ۳۲ (۱): ۱۷–۸۲. doi:10.1006/jhev.1996.0097. PMID 9034954.
- Bill Bryson (2004). "28. The Mysterious Biped". A Short History of Nearly Everything. Random House, Inc. pp. ۵۲۲–۵۴۳. ISBN 978-0-385-66004-4.
- Ungar, Peter; Teaford, Mark F. (2002), Human Diet: Its Origin and Evolution, Bergin & Garvey, p. ۲۰۶, ISBN 978-0-89789-736-5
- Bogin, Barry (1997). "The evolution of human nutrition". The Anthropology of Medicine: From Culture to Method. Bergen and Garvey. p. ۹۶–۱۴۲. ISBN 978-0-89789-516-3.
- "Human nutrition: evolutionary perspectives". Integr Physiol Behav Sci. ۴۰ (۲): ۱۱۴–۱۷. April/June 2005. doi:10.1007/BF02734246. PMID 17393680. Unknown parameter
|نام خانوداگی=ignored (help);|first1=missing|last1=in Authors list (help); Check date values in:|تاریخ=(help) - Robertson, ML; Snodgrass, JJ (2007), "Effects of brain evolution on human nutrition and metabolism", Annu Rev Nutr., ۲۷, p. ۳۱۱–۲۷
- Wood, B. & Collard, M. (1999) The changing face of Genus Homo. Evol. Anth. 8(6) 195-207
- Toothy Tree-Swinger May Be Earliest Human, News.discovery.com, 2010-05-21
- Wood B (1999). "'Homo rudolfensis' Alexeev, 1986-fact or phantom?". J. Hum. Evol. ۳۶ (۱): ۱۱۵–۸. doi:10.1006/jhev.1998.0246. PMID 9924136.
- Gabounia L. de Lumley M. Vekua A. Lordkipanidze D. de Lumley H. (2002). "Discovery of a new hominid at Dmanisi (Transcaucasia, Georgia)". Comptes Rendus Palevol. ۱ (۴): ۲۴۳–۵۳. doi:10.1016/S1631-0683(02)00032-5.
- Lordkipanidze D, Vekua A, Ferring R, et al. (2006). "A fourth hominin skull from Dmanisi, Georgia". The anatomical record. Part A, Discoveries in molecular, cellular, and evolutionary biology. ۲۸۸ (۱۱): ۱۱۴۶–۵۷. doi:10.1002/ar.a.20379. PMID 17031841.
- Turner W (1895). "On M. Dubois' Description of Remains recently found in Java, named by him Pithecanthropus erectus: With Remarks on so-called Transitional Forms between Apes and Man". Journal of anatomy and physiology. ۲۹ (Pt 3): ۴۲۴–۴۵. PMID 17232143.
- Spoor F, Wood B, Zonneveld F (1994). "Implications of early hominid labyrinthine morphology for evolution of human bipedal locomotion". Nature. ۳۶۹ (۶۴۸۲): ۶۴۵–۸. doi:10.1038/369645a0. PMID 8208290.
- Bermúdez de Castro JM, Arsuaga JL, Carbonell E, Rosas A, Martínez I, Mosquera M (1997). "A hominid from the lower Pleistocene of Atapuerca, Spain: possible ancestor to Neandertals and modern humans". Science. ۲۷۶ (۵۳۱۷): ۱۳۹۲–۵. doi:10.1126/science.276.5317.1392. PMID 9162001.
- Carbonell, Eudald (2008-03-27). "The first hominin of Europe". Nature. ۴۵۲ (۷۱۸۶): ۴۶۵–۴۶۹. doi:10.1038/nature06815. PMID 18368116. Archived from the original on 22 November 2010. Retrieved 2008-03-26. Unknown parameter
|coauthors=ignored (|author=suggested) (help) - Manzi G, Mallegni F, Ascenzi A (2001). "A cranium for the earliest Europeans: Phylogenetic position of the hominid from Ceprano, Italy". Proc. Natl. Acad. Sci. U.S.A. ۹۸ (۱۷): ۱۰۰۱۱–۶. doi:10.1073/pnas.151259998. PMID 11504953.
- Czarnetzki, A; Jakob, T; Pusch, CM (2003). "Palaeopathological and variant conditions of the Homo heidelbergensis type specimen (Mauer, Germany)". Journal of Human Evolution. ۴۴ (۴): ۴۷۹. doi:10.1016/S0047-2484(03)00029-0. PMID 12727464. More than one of
|pages=and|page=specified (help) - "Scientists discover hominid cranium in Ethiopia" (Press release). Indiana University. March 27, 2006. Retrieved ۲۰۰۶-۱۱-۲۶.
- Harvati K (2003). "The Neanderthal taxonomic position: models of intra- and inter-specific craniofacial variation". J. Hum. Evol. ۴۴ (۱): ۱۰۷–۳۲. doi:10.1016/S0047-2484(02)00208-7. PMID 12604307.
- Herrera, K. J. ; Somarelli, J. A. ; Lowery, R. K. ; Herrera, R. J. (2009). "To what extent did Neanderthals and modern humans interact?". Biological Reviews 84 (2): 245–257. DOI:10.1111/j.1469-185X.2008.00071.x. PMID 19391204
- Krings M, Stone A, Schmitz RW, Krainitzki H, Stoneking M, Pääbo S (1997). "Neandertal DNA sequences and the origin of modern humans". Cell. ۹۰ (۱): ۱۹–۳۰. doi:10.1016/S0092-8674(00)80310-4. PMID 9230299.
- Green RE, et al. (2008). "A complete Neandertal mitochondrial genome sequence determined by high-throughput sequencing". Cell. ۱۳۴ (۳): ۴۱۶–۴۲۶. doi:10.1016/j.cell.2008.06.021. PMID 18692465.
- Serre D, Langaney A, Chech M, et al. (2004). "No Evidence of Neandertal mtDNA Contribution to Early Modern Humans". PLoS Biol. ۲ (۳): E57. doi:10.1371/journal.pbio.0020057. PMID 15024415.
- Jennifer Viegas (2010-05-06). "Neanderthals, humans interbred, DNA proves". Discovery News. Archived from the original on 22 November 2010. Retrieved 2010-08-17. External link in
|publisher=(help) - Gutiérrez G, Sánchez D, Marín A (2002). "A reanalysis of the ancient mitochondrial DNA sequences recovered from Neandertal bones". Mol. Biol. Evol. ۱۹ (۸): ۱۳۵۹–۶۶. PMID 12140248.
- Hebsgaard MB, Wiuf C, Gilbert MT, Glenner H, Willerslev E (2007). "Evaluating Neanderthal genetics and phylogeny". J. Mol. Evol. ۶۴ (۱): ۵۰–۶۰. doi:10.1007/s00239-006-0017-y. PMID 17146600.
- Diamond, Jared (1992), The Third Chimpanzee: The Evolution and Future of the Human Animal, Harper Perennial, ISBN 978-0-06-098403-8
- How Neanderthals met a grisly fate: devoured by humans. The Observer. May 17, 2009
- "DNA identifies new ancient human dubbed 'X-woman'". BBC News. March 25, 2010
- Sample, Ian (March 24, 2010), New species of human ancestor found in Siberia, The Guardian, archived from the original on ۲۰۱۰-۱۱-۲۲
- Krause, Johannes; Fu, Qiaomei; Good, Jeffrey M.; Viola, Bence; Shunkov, Michael V.; Derevianko, Anatoli P. & Pääbo, Svante (2010). "The complete mitochondrial DNA genome of an unknown hominin from southern Siberia". Nature. ۴۶۴ (۷۲۹۰): ۸۹۴–۸۹۷. doi:10.1038/nature08976. PMID 20336068.
- Katsnelson, Alla (March 24, 2010), New hominin found via mtDNA, The Scientist, archived from the original on ۲۰۱۰-۱۱-۲۲
- "Denisova Admixture and the First Modern Human Dispersals into Southeast Asia and Oceania". The American Journal of Human Genetics. ۸۹ (۱۴). doi:10.1016/j.ajhg.2011.09.005. PMC 3188841. PMID 21944045.
- Brown P, Sutikna T, Morwood MJ, et al. (2004). "A new small-bodied hominin from the Late Pleistocene of Flores, Indonesia". Nature. ۴۳۱ (۷۰۱۲): ۱۰۵۵–۶۱. doi:10.1038/nature02999. PMID 15514638.
- Argue, D; Donlon, D; Groves, C; Wright, R (2006), "Homo floresiensis: microcephalic, pygmoid, Australopithecus, or Homo?", J. Hum. Evol., ۵۱ (۴), p. ۳۶۰–۷۴, doi:10.1016/j.jhevol.2006.04.013, PMID 16919706
- Martin, RD; Maclarnon, AM; Phillips, JL; Dobyns, WB (2006), "Flores hominid: new species or microcephalic dwarf?", The anatomical record. Part A, Discoveries in molecular, cellular, and evolutionary biology, ۲۸۸ (۱۱), p. ۱۱۲۳–۴۵, doi:10.1002/ar.a.20389, PMID 17031806
- Ambrose, Stanley H. (1998). "Late Pleistocene human population bottlenecks, volcanic winter, and differentiation of modern humans". Journal of Human Evolution. ۳۴ (۶): ۶۲۳–۶۵۱. doi:10.1006/jhev.1998.0219. PMID 9650103.
- Ambrose, Stanley H. (2005), Volcanic Winter, and Differentiation of Modern Humans, Bradshaw Foundation
- Gibbons, Ann (1998). "Solving the Brain's Energy Crisis". Science. ۲۸۰ (۵۳۶۸): ۱۳۴۵–۴۷. doi:10.1126/science.280.5368.1345. PMID 9634409.
- Howell, F.; Bourlière, François, African Ecology and Human Evolution, Transaction Publishers, p. ۳۹۸, ISBN 978-0-202-36136-9
- Freeman, Scott; Herron, Jon C. (2007), Evolutionary Analysis, Pearson Education, Inc., p. ۷۸۶–۷۸۸, ISBN 978-0-13-227584-2
- Plummer, T (2004), "Flaked stones and old bones: Biological and cultural evolution at the dawn of technology", Am. J. Phys. Anthropol., Suppl 39, p. ۱۱۸–۶۴, doi:10.1002/ajpa.20157, PMID 15605391
- Ambrose, SH (2001), "Paleolithic technology and human evolution", Science, ۲۹۱ (۵۵۰۹), p. ۱۷۴۸–۵۳, doi:10.1126/science.1059487, PMID 11249821
- Mcbrearty, S; Brooks, A. S. (2000), "The revolution that wasn't: a new interpretation of the origin of modern human behavior", J. Hum. Evol., ۳۹ (۵), p. ۴۵۳–۵۶۳, doi:10.1006/jhev.2000.0435, PMID 11102266
- Figure 1. Phylogeny of Primate Lice from Morphological and Molecular Data, in Clayton, DH; Smith, VS; Hammond, SL; Rogers, AR (2004). "Genetic Analysis of Lice Supports Direct Contact between Modern and Archaic Humans". PLoS Biol. ۲ (۱۱): e340. doi:10.1371/journal.pbio.0020340. PMC 521174. PMID 15502871. Archived from the original on ۲۰۱۰-۱۱-۲۲.
- Modern Humans Came Out of Africa, "Definitive" Study Says, News.nationalgeographic.com, 2010-10-28
- Stringer, CB; Andrews, P (1988), "Genetic and fossil evidence for the origin of modern humans", Science, ۲۳۹ (۴۸۴۵), p. ۱۲۶۳–۸, doi:10.1126/science.3125610, PMID 3125610 Unknown parameter
|ماه=ignored (help) - Cann, RL; Stoneking, M; Wilson, AC (1987), "Mitochondrial DNA and human evolution", Nature, ۳۲۵ (۶۰۹۹), p. ۳۱–۶, doi:10.1038/325031a0, PMID 3025745, archived from the original on ۲۰۱۰-۱۱-۲۲
- Behar et al. 2008, Gonder et al. 2007, Reed and Tishkoff
- Searching for traces of the Southern Dispersal بایگانیشده در ۱۰ مه ۲۰۱۲ توسط Wayback Machine, by Dr. Marta Mirazón Lahr, et al.
- Hill, C; Achilli, A; Rengo, C; Clarke, D; Meehan, W; Blackburn, J; Semino, O; Scozzari, R (2005). "Single, Rapid Coastal Settlement of Asia Revealed by Analysis of Complete Mitochondrial Genomes". Science. ۳۰۸ (۵۷۲۴): ۱۰۳۴–۶. doi:10.1126/science.1109792. PMID 15890885.
|first1=missing|last1=in Authors list (help) - Gill, Victoria (May 1, 2009), "Africa's genetic secrets unlocked", BBC News
- "The new batch - 150,000 years ago". BBC - Science & Nature - The evolution of man.
- When humans faced extinction, BBC, 2003-06-09, archived from the original on ۲۰۱۰-۱۱-۲۲
- Wolpoff, MH (2000). "Multiregional, not multiple origins". Am J Phys Anthropol. ۱۱۲ (۱): ۱۲۹–۳۶. doi:10.1002/(SICI)1096-8644(200005)112:1<129::AID-AJPA11>3.0.CO;2-K. PMID 10766948.
- Wolpoff, MH (1988). "Modern Human Origins". Science. ۲۴۱ (۴۸۶۷): ۷۷۲–۴. doi:10.1126/science.3136545. PMID 3136545.
- Leakey, Richard (1994). The Origin of Humankind. Basic Books. p. ۸۷–۸۹. ISBN 978-0-465-05313-1.
- Jorde, LB; Bamshad, M; Rogers, AR (1998). "Using mitochondrial and nuclear DNA markers to reconstruct human evolution". BioEssays. ۲۰ (۲): ۱۲۶–۳۶. doi:10.1002/(SICI)1521-1878(199802)20:2<126::AID-BIES5>3.0.CO;2-R. PMID 9631658. Unknown parameter
|ماه=ignored (help) - Wall, J. D.; Lohmueller, K. E.; Plagnol, V. (2009). "Detecting Ancient Admixture and Estimating Demographic Parameters in Multiple Human Populations". Molecular Biology and Evolution. ۲۶ (۸): ۱۸۲۳–۷. doi:10.1093/molbev/msp096. PMC 2734152. PMID 19420049.
- Green RE, Krause J, Briggs AW, et al. (2010). "A draft sequence of the Neandertal genome". Science. ۳۲۸ (۵۹۷۹): ۷۱۰–۲۲. doi:10.1126/science.1188021. PMID 20448178. Unknown parameter
|ماه=ignored (help) - ^ Reich D, Green RE, Kircher M, et al. (December 2010). "Genetic history of an archaic hominin group from Denisova Cave in Siberia". Nature 468 (7327): 1053–60. doi:10.1038/nature09710. PMID 21179161
- Reich D, Patterson N, Kircher M, et al. (2011). "Denisova admixture and the first modern human dispersals into Southeast Asia and Oceania". Am. J. Hum. Genet. ۸۹ (۴): ۵۱۶–۲۸. doi:10.1016/j.ajhg.2011.09.005. PMC 3188841. PMID 21944045. Unknown parameter
|ماه=ignored (help) - Byars, S. G. ; Ewbank, D. ; Govindaraju, D. R. ; Stearns, S. C. (2009). "Natural selection in a contemporary human population". Proceedings of the National Academy of Sciences 107 (suppl_1): 1787–1792. Bibcode 2010PNAS..107.1787B. DOI:10.1073/pnas.0906199106. PMID 19858476.
- Cochran G and Harpending H. 2009. The 10,000 Year Explosion. Basic Books N.Y.
- Richard Dawkins, The magic of reality: how we know what's real, fP free Press, pages 32-53
- طباطبایی، تفسیر المیزان، ج۴، سوره نساء، آیه ۱، ص ۲۱۲ _ ۲۳۸
- طباطبایی، تفسیر المیزان، ج۴، سوره نساء، تفسیر آیه ۱، ص ۲۱۲ الی ۲۳۸
- طباطبایی، ترجمهالمیزان، ج۴، ص ۲۳۱
منابع برای مطالعهٔ بیشتر
- Alexander, R. D. (1990). "How did humans evolve? Reflections on the uniquely unique species" (PDF). University of Michigan Museum of Zoology Special Publication. University of Michigan Museum of Zoology (۱): ۱–۳۸. Archived from the original (PDF) on 11 April 2012. Retrieved 14 June 2012.
- Enard, Wolfgang; et al. (2002-08-22). "Molecular evolution of FOXP2, a gene involved in speech and language". Nature. ۴۱۸ (۶۹۰۰): ۸۶۹–۷۲ [۸۷۰]. doi:10.1038/nature01025. PMID 12192408.
- Flinn, M. V. , Geary, D. C. , & Ward, C. V. (2005). Ecological dominance, social competition, and coalitionary arms races: Why humans evolved extraordinary intelligence. Evolution and Human Behavior, 26, 10-46. Full text.پیدیاف (345 KB)
- Geary, David C (2005), The Origin of Mind: Evolution of Brain, Cognition and General Intelligence, Washington DC: American Psychological Association, ISBN 1-59147-181-8
- Green, Richard E; et al. (7 May 2010), "A Draft Sequence of the Neandertal Genome", Science, ۳۲۸ (۵۹۷۹), p. ۷۱۰–۷۲۲
- Gibbons, Ann. The First Human: The Race to Discover our Earliest Ancestor. Anchor Books (2007). ISBN 978-1-4000-7696-3
- Hartwig, Walter, ed. (2002. Reprinted 2004). The Primate Fossil Record. Cambridge University Press. ISBN 978-0-521-08141-2. Check date values in:
|year=(help). - Heizmann, Elmar P J, Begun, David R (2001). "The oldest Eurasian hominoid". Journal of Human Evolution. ۴۱ (۵): ۴۶۳. doi:10.1006/jhev.2001.0495. PMID 11681862. More than one of
|pages=and|page=specified (help) - Hill, Andrew; Ward, Steven (1988). "Origin of the hominidae: The record of African large hominoid evolution between 14 my and 4 my". Yearbook of Physical Anthropology. ۳۱ (۵۹): ۴۹–۸۳. doi:10.1002/ajpa.1330310505.
- Ijdo,J.W; Baldini, A; Ward,D.C; Reeders,S.T; Wells, R.A (1991). "Origin of human chromosome 2: An ancestral telomere-telomere fusion" (PDF). Genetics. ۸۸ (۲۰): ۹۰۵۱–۹۰۵۵. Unknown parameter
|month=ignored (help)—two ancestral ape chromosomes fused to give rise to human chromosome 2 - Johanson, Donald & Wong, Kate. Lucy's Legacy: The Quest for Human Origins. Three Rivers Press (2009). ISBN 978-0-307-39640-2
- Jones, Steve; Martin, Robert D. ; Pilbeam, David R (Editors). (1994). The Cambridge Encyclopedia of Human evolution. Cambridge University Press. ISBN 978-0-521-46786-5. (Note: this book contains very useful, information dense chapters on primate evolution in general, and human evolution in particular, including fossil history).
- Leakey, Richard & Lewin, Roger. Origins Reconsidered: In Search of What Makes us Human. Little, Brown and Company (1992). ISBN 0-316-90298-5
- Lewin, Roger (2005), Human Evolution: An Illustrated Introduction (5th ed ed.), Blackwell Publishing, ISBN 1-4051-0378-7
- Lewin, Roger. Bones of Contention: Controversies in the Search for Human Origins. Penguin Books (1987). ISBN 0-14-022638-9
- Morwood, Mike & van Oosterzee, Penny. A New Human: The Startling Discovery and Strange Story of the 'Hobbits' of Flores, Indonesia. Smithsonian Books (2007). ISBN 978-0-06-089908-0
- Oppenheimer, Stephen. Out of Eden: The Peopling of the World. Constable (2003). ISBN 1-84119-697-5
- Ovchinnikov; Götherström, Anders; Romanova, Galina P.; Kharitonov, Vitaliy M.; Lidén, Kerstin; Goodwin, William; et al. (2000). "Molecular analysis of Neanderthal DNA from the Northern Caucasus". Nature. ۴۰۴ (۶۷۷۷): ۴۹۰. doi:10.1038/35006625. PMID 10761915. More than one of
|pages=and|page=specified (help) - Roberts, Alice. The Incredible Human Journey: The Story of how we Colonised the Planet. Bloomsbury (2009). ISBN 978-0-7475-9839-8
- Sarmiento, Esteban et al. The Last Human: A guide to twenty-two species of extinct humans. New Haven and London: Yale University Press, 2007. ISBN 978-0-300-10047-1
- Shreeve, James. The Neanderthal Enigma: Solving the Mystery of Modern Human Origins. Viking (1996). ISBN 0-670-86638-5
- Stringer, Chris. The Origin of Our Species. Allen Lane (2011). ISBN 978-1-84614-140-9
- Stringer, Chris & Andrews, Peter. The Complete World of Human Evolution. Thames & Hudson (2005). ISBN 0-500-05132-1
- Stringer, Chris & McKie, Robin. African Exodus: The Origins of Modern Humanity. Jonathan Cape (1996). ISBN 0-224-03771-4
- van Oosterzee, Penny. The Story of Peking Man. Allen & Unwin (1999). ISBN 1-86508-632-0
- Walker, Allan & Shipman, Pat. The Wisdom of the Bones: In Search of Human Origins. Weidenfeld and Nicholson (1996). ISBN 0-297-81670-5
- Wade, Nicholas. Before the Dawn: Recovering the Lost History of our Ancestors. Penguin Press (2006). ISBN 978-0-7156-3658-9
- Weiss, M.L. , & Mann, A.E (1985). "'Human Biology and Behaviour: An anthropological perspective" (4th ed.). Boston: Little Brown. ISBN 978-0-673-39013-4. (Note: this book contains very accessible descriptions of human and non-human primates, their evolution, and fossil history).
- Wells, Spencer. The Journey of Man: A Genetic Odyssey. Allen Lane/The Penguin Press (2002). ISBN 0-7139-9625-0
- Wood, Bernard and Mark Collard, "The Human Genus", Science, New Series, ۲۸۴ (۵۴۱۱), p. ۶۵–۷۱
پیوند به بیرون
| در ویکیانبار پروندههایی دربارهٔ فرگشت انسان موجود است. |
- BBC: The Evolution of Man
- Illustrations from Evolution (textbook)
- Smithsonian – Homosapiens
- Smithsonian – The Human Origins Program
- Becoming Human: Paleoanthropology, Evolution and Human Origins, presented by Arizona State University's Institute of Human Origins
- species
- Bones, Stones and Genes: The Origin of Modern Humans, Howard Hughes Medical Institute 2011 Holiday Lecture Series.
- DNA Shows Neandertals Were Not Our Ancestors بایگانیشده در ۲۰۰۵-۱۰-۲۵ توسط Wayback Machine
- BBC: Finds test human origins theory. 2007-08-08 Homo habilis and Homo erectus are sister species that overlapped in time.
- „Der Ursprung des Menschen: Auf den Spuren der Hominiden. “ – Dossier von spektrumdirekt vom 12. Januar 2010
- „Die Entwicklungsgeschichte des menschlichen Stammbaums“ – Umfangreiche Übersichtsarbeit
- „Evolution des Menschen. “ – Film vom 16. Juli 2008 in der ZDFmediathek
| طبقهبندی (انسانتباران) |
| ||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| مدلها |
| ||||||||||||||||||||||||||
| خط زمانی |
| ||||||||||||||||||||||||||
| دیگر |
| ||||||||||||||||||||||||||
مقالات مرتبط کپیها | ||
|---|---|---|
| گونههای کپی |
| |
| مطالعه کپی |
| |
| وضعیت حقوقی و اجتماعی |
| |
| همچنین نگاه کنید به |
| |
| ||
| فرگشت |
|  |
| ژنتیک جمعیت | ||
| تکوین |
| |
| آرایه |
| |
| عضو |
| |
| فرآیند |
| |
| تمپوها و مدها |
| |
| گونهزایی |
| |
| تاریخچه اندیشه فرگشتی |
| |
| فلسفه |
| |
| مرتبط |
| |
| ||