دگم مرکزی زیستشناسی مولکولی
دگم یا باور مرکزی زیستشناسی مولکولی (به انگلیسی: Central Dogma of Molecular Biology) توضیحی برای جریان اطلاعات ژنتیکی در یک سیستم بیولوژیکی است که اغلب به این صورت است DNA باعث ساخت RNA شده و RNA پروتئین را میسازد[1] اگر چه این معنی متفاوت از معنای ابتدایی آن است. این ایده اولین بار توسط فرانسیس کریک در سال ۱۹۵۸ مطرح شد:[2]
| « | The Central Dogma. This states that once 'information' has passed into پروتئین it cannot get out again. In more detail, the transfer of information from نوکلوئیک اسید to nucleic acid, or from nucleic acid to protein may be possible, but transfer from protein to protein, or from protein to nucleic acid is impossible. Information means here the precise determination of sequence, either of bases in the nucleic acid or of amino acid residues in the protein. | » |
—Francis Crick, 1958 | ||
و در مجله Nature در سال ۱۹۷۰ بدین صورت دوباره بیان شده:[3]
| « | The central dogma of molecular biology deals with the detailed residue-by-residue transfer of sequential information. It states that such information cannot be transferred back from protein to either protein or nucleic acid. | » |
—Francis Crick | ||
نسخه دومی از دگم مرکزی وجود دارد که علیرغم مقبولیت بیشتر، صحیح نیست. مسیر ساده شدهٔ DNA → RNA → پروتئین است که توسط جیمز واتسون در اولین نسخه زیستشناسی مولکولی ژن (۱۹۶۵) منتشر شد. نسخه واتسون به دلیل توصیف دو مرحله ای مسیر (DNA → RNA و RNA → پروتئین) به عنوان قاعده مرکزی متفاوت از نظریه کریک است. قاعده ارائه شده توسط کریک هنوز معتبر است درحالیکه نسخه واتسون اینطور نیست.
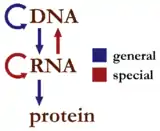
این قاعده چارچوبی برای درک انتقال توالی اطلاعات بین حاملهای اطلاعاتی زیست بسپار، در معنای عمومی، در موجودات زنده است. ۳ طبقه عمده این زیست بسپارها: DNA و RNA (اسیدهای نوکلئیک) و پروتئین هستند. ۳×۳=۹, صورت قابل تصور از انتقال مستقیم اطلاعات میتواند در این میان رخ دهد. بر اساس قاعده این سه دسته به سه گروه تقسیم میشوند: سه انتقال عمومی (باور بر این است که بهطور معمول در اکثر سلولها رخ میدهد)، سه انتقال ویژه (به صورت معمول تنها در شرایط خاص در مورد برخی از ویروسها یا در آزمایشگاه اتفاق میافتد) و سه انتقال ناشناخته (اعتقاد بر این است که هرگز رخ نمیدهد). بهطور کلی انتقال عمومی جریان طبیعی اطلاعات زیستی را توصیف میکند: DNA به DNA بازنویسی میشود (DNAرونویسی) اطلاعات DNA همچنین میتواند روی mRNA (رونویسی) بازنویسی شده و با ترجمه اطلاعات موجود در mRNA به عنوان یک الگو (ترجمه) پروتئینها ساخته میشوند. انتقال ویژه بدین صورت است که: RNA از روی RNA کپی شده (RNA رونویسی) و DNA با استفاده از RNA الگو (رونویسی معکوس) سنتز شده و در نهایت پروتئین مستقیماً و بدون استفاده از mRNA از روی DNA الگو ساخته میشود. انتقال ناشناخته اینگونه است که: یک پروتئین که از یک پروتئین بازنویسی شده، با استفاده از ساختار اولیهٔ پروتئین به عنوان RNA الگو را ساخته و DNA نیز با استفاده از ساختار اولیهٔ یک پروتئین به عنوان یک الگو - ساخته میشود، که البته بهطور طبیعی رخ نمیدهد.
توالی اطلاعات زیستی
زیست بسپارهایی که از DNA,RNA و (پلی) پپتیدها ساخته شدهاند پلیمرهای خطی هستند (یعنی هر مونومر یه حداکثر دو مونومر دیگر متصل است). توالی مونومرها به شکل مؤثری اطلاعات را کدگذاری میکند. انتقال اطلاعات تحت دگم مرکزی به شکل مؤثری پایدار بوده و انتقال قطعی است (برای وقتی است که توالی زیست بسپاری به عنوان الگو برای تولید زیست بسپار دیگر با توالی وابسته به توالی زیست بسپار اصلی مورد استفاده قرار میگیرد)
انتقال عمومی اطلاعات توالیهای زیستی
جدول سه دسته انتقال اطلاعات تحت نظام قاعده General Special Unknown DNA → DNA RNA → DNA protein → DNA DNA → RNA RNA → RNA protein → RNA RNA → protein DNA → protein protein → protein
همانندسازی DNA
در این معنا، رونویسی DNA در صورت فراهم بودن مواد ژنتیکی مورد نیاز برای اخلاف سلول، پیکری یا تولیدمثلی، انجام میشود و بازنویسی از DNA به DNA مرحله اساسی در دگم مرکزی بهشمار میرود. گروهی پیچیده از پروتئینها به نام رپلیزوم Replisome رونویسی از اطلاعات رشته والدی را به رشته مکمل دختری انجام میدهند.
رپلیزوم تشکیل شدهاست از:
- یک هلیکاز که ابرمارپیچ را مثل مارپیچ دو رشتهای DNA باز میکند تا چنگال رونویسی بسازد. پروتئین SSB که میچسبد تا رشته دو تایی DNA را باز نگه دارد و از پیوستگی مجدد آن جلوگیری کند. RNAپرایماز که پرایمر مکمل RNA را به هر رشته الگو چسبانده تا رونویسی شروع شود. DNAپلیمراز۲ که رشتهٔ الگوی موجود را از سمت ۳' به ۵' میخواند و نوکلئوتیدهای مکمل را از انتهای ۵' به انتهای ۳' رشتهٔ دختری اضافه میکند.
این فرایند در مرحله S چرخه سلولی اتفاق میافتد.
رونویسی
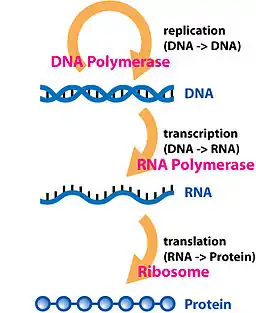
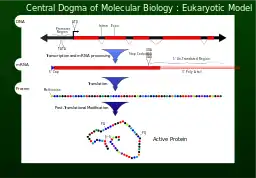
رونویسی روندی است که اطلاعات موجود در یک بخش از DNA در قالب یک قطعه تازه سر هم شده RNA پیامبر messenger RNA (mRNA) رونویسی میشوند. آنزیمهایی که این فرایند را تسهیل میکنند شامل RNA پلیمراز و عوامل رونویسی هستند. در سلولهای یوکاریوت سلولهای اولیه نسخه اولیه pre-mRNA. است Pre-mRNA باید برای ترجمه مورد پردازش قرار بگیرد. پردازش شامل اضافه شدن یک کلاه در سمت ۵' و یک دم پلی-A به زنجیره pre-mRNA است که پس از پیرایشُ Splicing اتفاق میافتد..پیرایش زمانی اتفاق میافتد که میزان مناسب و در حال افزایش تنوع پروتئینی میتواند سبب تولید یک نوع mRNA شود. این محصول از کل فرایند رونویسی (که با تولید زنجیرههای pre-mRNA آغاز شدهاست), یک زنجیره mRNA بالغ را تشکیل میدهد.
ترجمه
mRNA بالغ برای فرایند ترجمه به ریبوزوم میروند. در سلولهای پروکاریوتی که بخشبندی تحت عنوان هسته ندارند، فرایند رونویسی و ترجمه بدون جدایی مشخص و به صورت پیوسته اتفاق میافتد. در سلولهای یوکاریوتی، محل رونویسی (هسته سلول) معمولاً از محل ترجمه (سیتوپلاسم) جداست، بنابراین mRNA باید از هسته به سیتوپلاسم که محل قرارگیری ریبوزوم است برده شود. ریبوزوم کدهای ژنتیکی (Codon) 3تایی mRNA را که معمولاً با AUG (آدنین، یوراسیل، گوانین)، یا کدون شروعکننده متیونین در پایین دست downstream محل اتصال ریبوزوم شروع میشود را میخواند. مجموعه فاکتورهای شروعکننده و فاکتورهای طویلکننده RNAهای انتقالی آمینواسیله شده (tRNA) را به مجموعه mRNA ریبوزوم آورده و کدونهای mRNA را با آنتی کدونهای tRNA جور میکنند. هر tRNA رزیدوی مناسب را برای تولید زنجیره پلی پپتیدی مورد سنتز حمل میکند. با اتصال اسیدهای آمینه به رشته پپتیدی در حال رشد، زنجیره شروع به پیچ خوردن به شکل صحیح فضایی خود میکند. فرایند ترجمه به کدون خاتمه که به شکلهای UAA،UGAو یا UAG هستند پایان میپذیرد.
mRNA تمام اطلاعات لازم برای تخصصی کردن طبیعت پروتئین بالغ را ندارد. زنجیره پلی پپتیدی در حال شکلگیری که از ریبوزوم رها میشود پردازشهای بیشتری را قبل از آزاد شدن نهایی محصول نیاز دارد. مثلاً برای تا خوردن صحیح که پیچیده و ضروری است، بیشتر پروتئینها به پروتئینهای چاپرون نیاز دارند تا شکل صحیح محصول را کنترل نماید. برخی پروتئینها قصعاتی داخلی را از زنجیره پلی پپتیدی خود کرده و سمت آزاد خود را پیرایه میکنند. در این فرایندها، قطعات دور انداخته شده داخلی اینین Intein خوانده میشوند. سایر پروتئینها باید بدون پیرایش به قطعات چندگانه تقسیم شوند. برخی زنجیرههای پلی پپتیدی باید به صورت متقابل متصل شوند و برخی دیگر باید به کوفاکتورهایی مثل Haem (heme) متصل شوند تا قابلیت عملکردی پیدا کنند.
منابع
- Leavitt, Sarah A. (June 2010). "Deciphering the Genetic Code: Marshall Nirenberg". Office of NIH History. Archived from the original on 17 March 2015.
- Crick, F.H.C. (1958). "On Protein Synthesis". In F.K. Sanders. Symposia of the Society for Experimental Biology, Number XII: The Biological Replication of Macromolecules. Cambridge University Press. pp. 138–163.
- Crick, Francis (August 1970). "Central dogma of molecular biology" (PDF). Nature. 227 (5258): 561–3. Bibcode:1970Natur.227..561C. doi:10.1038/227561a0. PMID 4913914.
| نمای کلی |
| ||||||
|---|---|---|---|---|---|---|---|
| مهندسی |
| ||||||
| |||||||
