زالو
زالو نوعی کرم حلقوی است. زالو حیوانی از گروه کرمهای آبزی و از راستهٔ آروارهداران است. بدن جانور قابلیت انقباض و انبساط فراوان دارد، طول بدن بین ۳ تا ۲۵ سانتیمتر و عرض بدن حدود ۱ سانتیمتر است. در جلوی بدن یک بادکش وجود دارد که مرکب از ۲ حلقه است و دهان در این ناحیه واقع شدهاست و در انتهای بدن یک بادکش مدور مرکب از ۷ حلقه دارد که توسط این بادکش خود را به یک نقطه متصل میکند و نقطهٔ اتکایی است برای حرکت کردن.
| زالو | |
|---|---|
 | |
| یک زالو در چین | |
| آرایهشناسی | |
| فرمانرو: | جانوران |
| زیرفرمانرو: | هوپسزیان |
| بالاشاخه: | چرخزیان |
| شاخه: | کرم حلقوی |
| رده: | کمربندبهتنان |
| زیررده: | Hirudinea Lamarck, ۱۸۱۸ |
| Infraclasses | |
|
Acanthobdellidea | |
زالو ۳ فک دارد و هر فک ۱۰۰ دندان دارد؛ یعنی یک زالو حدود ۳۰۰ دندان فوقالعاده ریز دارد.
اکثر زالوها در اطراف آبهای شیرین زندگی میکنند، در حالیکه بعضی گونهها را میتوان در خاک[1] و همچنین اطراف دریاها یافت. شناختهشدهترین زالوها، مثل زالوهای دارویی که در زالودرمانی استفاده میشوند خونخوراک هستند و از خون مهرهداران و همولنف بیمهرگان تغذیه میکنند.[2]بههرحال بیشتر گونههای زالوها شکارچی هستند و در درجه اول با بلع دیگر بیمهرگان تغذیه میکنند. در حال حاضر تقریباً ۷۰۰ گونه از زالوها شناخته شدهاند، که ۱۰۰ تا دریایی، ۹۰ تا زمینی و بقیهٔ آنها از آرایهٔ آبهای شیرین هستند.[3]
از لحاظ تاریخی زالوها در پزشکی (زالودرمانی)، برای گرفتن خون از بیماران استفاده میشدهاند.[4] سابقهٔ عمل زالودرمانی به هندوستان و یونان قدیم بازمیگردد که تا قرن هیجدهم و نوزدهم در اروپا و آمریکای شمالی ادامه یافت. در دوران مدرن، از زالو در مراحلی مانند چسباندن مجدد اجزای بدن در جراحیهای ترمیمی و پلاستیک[5] و در آلمان برای درمان آرتروز استفاده میکنند.[6][7]
آناتومی و فیزیولوژی
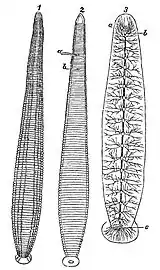
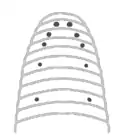
مثل دیگر حلقویتباران زالو حیوانی بندبند است، اما بیشباهت به دیگر حلقویتباران هیچ ارتباطی بین تقسیمبندی سطح خارجی بدن یک زالو و اندامهای داخلیاش وجود ندارد.[8] سطح بدن حیوان میتواند به ۱۰۲ حلقه تقسیم شود. درحالی که ساختار داخلیاش به ۳۲ بخش تقسیم شدهاست.[9] از ۳۲ بخش داخلی بدن، ۴ تای جلویی اختصاص به قسمت سر دارد، که شامل مغز قدامی و یک مکنده است. بعد از سر ۲۱ بند بعدی، بخش میانی بدن متشکل از ۲۱ غدهٔ عصبی، دو اندام تناسلی، و ۹ جفت بیضه است. سرانجام۷بند آخر ترکیب شدهاند تا دم و مغز خلفی جانور را تشکیل بدهند.
تولید مثل و نمو
زالوها هرمافرودیت هستند، به این معنی که هم اندام تناسلی ماده و هم نر را دارند (به ترتیب تخمدان و بیضه). زالوها با تکثیر متقابل تولیدمثل میکنند، و درهنگام آمیزش اسپرم منتقل میشود. زالوها مانند کرمهای خاکی، برای نگهداری تخمهایشان از کمربند تناسلی استفاده کرده و آنها را در پیلهای پنهان میکنند.
در حین آمیزش زالوها از تزریق زیرجلدی اسپرم خود استفاده میکنند. آنها از یک اسپرماتوفر، که یک ساختار حاوی اسپرم است، استفاده میکنند. گاهی در کنار یکدیگر، در حالی که جلوی یکی رو به پشت دیگری است قرار میگیرند و هر زالو اسپرماتوفر خود را به اندام دیگری تزریق میکند که اسپرمش از آنجا راه خود را به طرف اندام تناسلی زنانه پیدا خواهد کرد. رشد لارو به عنوان یک سری از مراحل رخ میدهد. در اولین مرحله سلول تقسیم شده و موجب به وجود آمدن یک جوانهسای AB و CD میشود و در مرحلهٔ تقسیم سلولی میانچهر است که سیتوپلاسم بدون یاخته که تلوپلاسم نامیده میشود شکل میگیرد.[10] تلوپلاسم به عنوان عامل تعیینکنندهٔ سرنوشت سلول D شناخته شدهاست.[11] در مرحلهٔ ۳ هنگام دومین تقسیم سلولی در بلاستومر CD تقسیمی نامساوی رخ میدهد. در نتیجه موجب به وجود آمدن یک سلول بزرگتر D در سمت چپ و یک سلول کوچکتر C در سمت راست میشود. این مرحلهٔ تقسیم نامساوی به تارچه،[12]بستگی دارد و در پایان مرحلهٔ ۳ سلول AB تقسیم میشود. در مرحلهٔ ۴ نمو، سلولهای بنیادی میکرومرها و تلوبلاستها تشکیل میشوند و در نتیجه، D quadrant برای تشکیل سلولهای پیشساز DM و DNOPQ تلوبلاست تقسیم میشود. در پایان مرحلهٔ ۶ زیگوت شامل ۲۵ میکرومر، ۳ ماکرومر(A, B وC)و ۱۰ تلوبلاست مشتق از D quadrant است.[13]
تلوبلاستها جفتهایی از پنج نوع مختلف (M, N, O, P, and Q) از یاخته بنیادی جنینی هستند که ستونهای بندبند سلولها را در سطح بدن جنین شکل میدهند.[14] یاختههای مشتق شده از تلوبلاست نوع M یاختههای مزودرم و مجموعهٔ کوچکی از یاختههای عصبی را میسازد، یاختههای مشتق شده از تلوبلاست نوع N منجر به ساخت بافتهای عصبی و تعدادی از اکتودرم شکمی میشود.Q در ساخت اکتودرم پشتی همکاری میکند و در زالوها O, P هم اثر هستند (دارای پتانسیل رشد یکسان) که اکتودرم جانبی تولید میکند. بههرحال تفاوت آن دو این است که P نسبت به O دسته بزرگتری از یاختههای روپوست پشتی و جانبی تولید میکند.[11]در کرمهای لجن برکه، بیشباهت به زالوها، تلو بلاستهای نوع O, P در همان مراحل اولیه نمو از هم مجزا میشوند و بنابراین یاختههای هم اثر نیستند.[15] هر بخش از بدن زالو از یک سلولاز نوع M, O، P و دو سلول از نوع N ,Q تشکیل شدهاست.[11]
اکتودرم و مزودرم تنه بهطور انحصاری از یاختههای تلوبلاست از ناحیهای به نام منطقه پیشرفت خلفی مشتق میشوند.[16][17] سر زالو که از ناحیهٔ بدون بندبند بیرون میآید، بوسیلهٔ اولین گروهی از میکرومرهایی که از یاختههای A, B، C, D مشتق شدهاند با ایجاد تقارن بین یاختههای AD و BC شکل میگیرد.
منابع
- مشارکتکنندگان ویکیپدیا. «Leech». در دانشنامهٔ ویکیپدیای انگلیسی، بازبینیشده در ۱۱ فوریه ۲۰۱۶.
- S. Fogden & J. Proctor; Proctor (1985). "Notes on the Feeding of Land Leeches (Haemadipsa zeylanica Moore and H. picta Moore) in Gunung Mulu National Park, Sarawak". Biotropica. 17 (2): 172–174. doi:10.2307/2388511. JSTOR 2388511.
- Roy Sawyer (1981). Kenneth, Muller; Nicholls, John; Stent, Gunther, eds. Neurobiology of the Leech. New York: Cold Spring Harbor Laboratory. pp. 7–26. ISBN 0-87969-146-8.
- Boris Sket, Peter Trontelj (2008). "Global diversity of leeches (Hirudinea) in freshwater". In E. V. Balian, C. Lévêque, H. Segers & K. Martens. Freshwater Animal Diversity Assessment. Hydrobiologia. 595. pp. 129–137. doi:10.1007/s10750-007-9010-8.
- Brian Payton (1981). Kenneth Muller, John Nicholls, & Gunther Stent, ed. Neurobiology of the Leech. New York: Cold Spring Harbor Laboratory. pp. 27–34. ISBN 0-87969-146-8.
- Adam, Robert; Zakrzewski, Peter (2001). "Therapeutic Use of Leeches: From the 'Annelids' or Medicine". 79 (1). University of Toronto Medical Journal: 65–7. Archived from the original on 19 December 2013. Retrieved 11 February 2016.
- M. Teut & A. Warning; Warning (2008). "Leeches, phytotherapy and physiotherapy in osteo-arthrosis of the knee—a geriatric case study". Forsch Komplementmed. 15 (5): 269–72. doi:10.1159/000158875. PMID 19001824.
- Michalsen, A; Moebus, S; Spahn, G; Esch, T; Langhorst, J; Dobos, GJ (2002). "Leech therapy for symptomatic treatment of knee osteoarthritis: Results and implications of a pilot study". Alternative therapies in health and medicine. 8 (5): 84–8. PMID 12233807.
- Ralph Buchsbaum, Mildred Buchsbaum, John Pearse, & Vicki Pearse (1987). Animals Without Backbone (3rd ed.). Chicago: The University of Chicago Press. pp. 312–317. ISBN 0-226-07874-4.
- Brian Payton (1981). Kenneth Muller, John Nicholls, & Gunther Stent, ed. Neurobiology of the Leech. New York: Cold Spring Harbor Laboratory. pp. 35–50. ISBN 0-87969-146-8.
- J. Fernandez, N. Olea, V. Tellez & C. Matte (1990). "Structure and development of the egg of the glossiphoniid leech Theromyzon rude: reorganization of the fertilized egg during completion of the first meiotic division". Developmental Biology. 137 (1): 142–154. doi:10.1016/0012-1606(90)90015-B. PMID 2295361.
- D. A. Weisblat & M. Shankland (1985). "Cell lineage and segmentation in the leech". Philosophical Transactions of the Royal Society B: Biological Sciences. 312 (1153): 39–56. doi:10.1098/rstb.1985.0176. JSTOR 2396301. PMID 2869529.
- D. C. Lyons & D. A. Weisblat (2009). "D quadrant specification in the leech Helobdella: actomyosin contractility controls the unequal cleavage of the CD blastomere". Developmental Biology. 334 (1): 46–58. doi:10.1016/j.ydbio.2009.07.007. PMC 3077801. PMID 19607823.
- M. Sandig & W. Dohle (1988). "The cleavage pattern in the leech Theromyzon tessulatum (Hirudinea, Glossiphoniidae)". Journal of Morphology. 196 (2): 217–252. doi:10.1002/jmor.1051960210. PMID 3385778.
- V. K. Berezovskii & M. Shankland (1996). "Segmental diversification of an identified leech neuron correlates with the segmental domain in which it expresses Lox2, a member of the Hox gene family". Journal of Neurobiology. 29 (3): 319–329. doi:10.1002/(SICI)1097-4695(199603)29:3<319::AID-NEU4>3.0.CO;2-C. PMID 8907161.
- A. Arai, A. Nakamoto & T. Shimizu (2001). "Specification of ectodermal teloblast lineages in embryos of the oligochaete annelid Tubifex: involvement of novel cell-cell interactions". Development. 128 (7): 1211–1219. PMID 11245587.
- D. Nardelli-Haefliger & M. Shankland (1993). "Lox10, a member of the NK-2 homeobox gene class, is expressed in a segmental pattern in the endoderm and in the cephalic nervous system of the leech Helobdella". Development. 118 (3): 877–892. PMID 7915671.
- M. Shankland & A. E. Bruce (1998). "Axial patterning in the leech: developmental mechanisms and evolutionary implications". Biological Bulletin. 195 (3): 370–372. doi:10.2307/1543150. JSTOR 1543150. PMID 9924777.
| در ویکیانبار پروندههایی دربارهٔ زالو موجود است. |
| بندپایان |  | |
|---|---|---|
| دیگر جانداران | ||
| بیماریهای منتقله | اسهال خونی • بیماری خواب • بیماری شاگاس • بیماری لایم • تب دنگی • تب خندق • تب راجعه شپشی • تب زرد • تب نیل غربی • تولارمی • تیفوس • تیفوس کنهای • سالک • طاعون • فیلاریاز • مالاریا • ورم ویروسی مغز • هاری | |
| جستارهای وابسته | ||